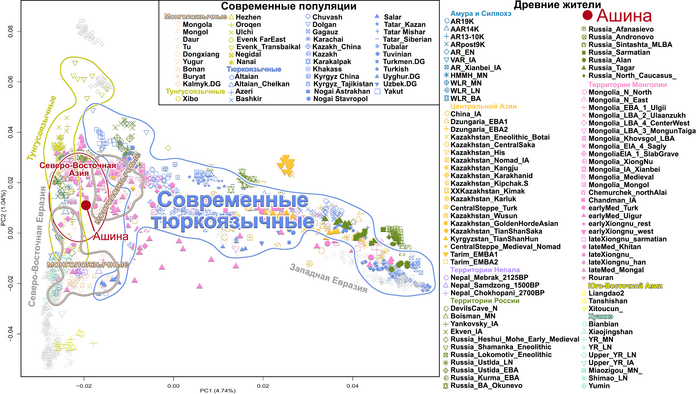
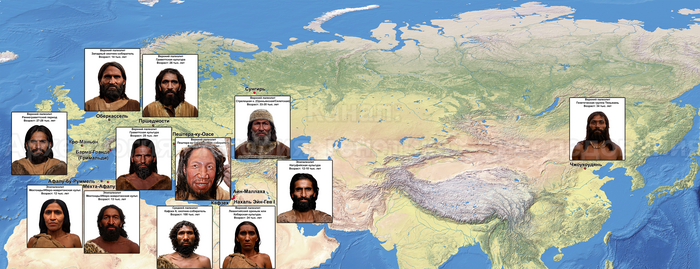
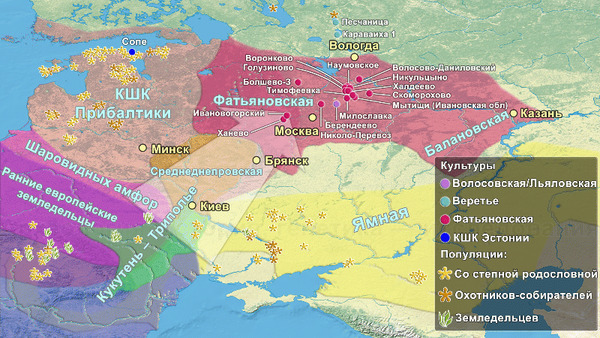
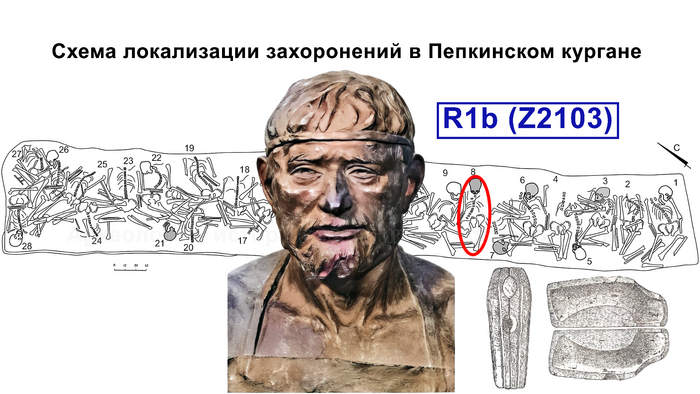
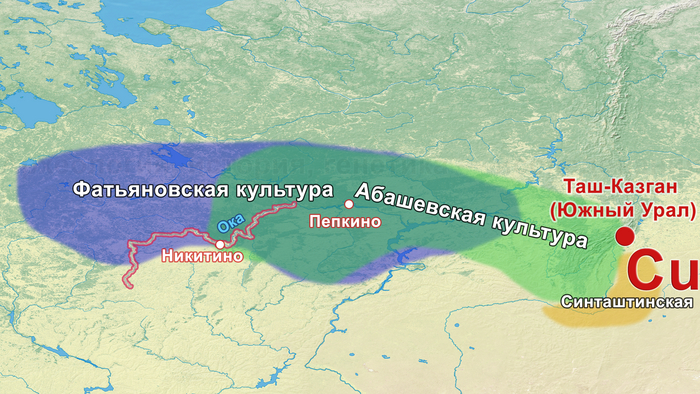
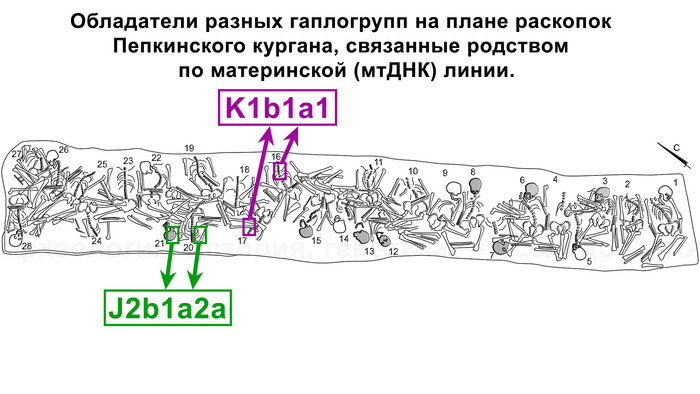
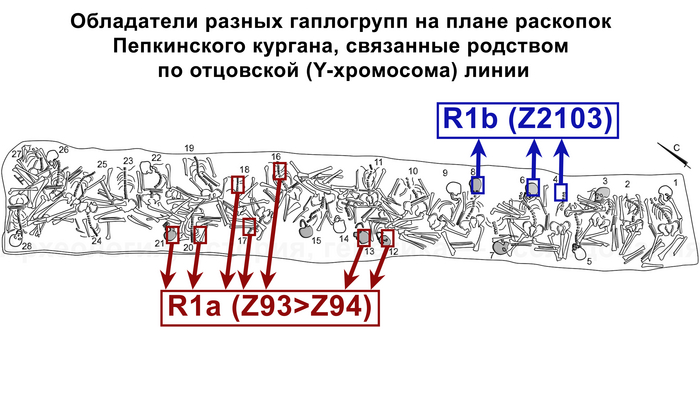
В статье речь пойдёт о генетических данных представительницы древних тюрков – принцессы, а после брака с императором китайско-сяньбийской династии Северной Чжоу - У-ди, императрицы – Ашины из одноимённого рода, дочери Мукан-кагана, третьего кагана древних тюрков – Ашина Сыцзинь, при котором Тюркский каганат расширил свои границы от Жёлтого до Чёрного моря, став крупнейшей империей того времени.
Кстати, о происхождении и внешности супруга Ашины - третьего императора китайско-сяньбийской династии Северной Чжоу - У-ди (Юйвэнь Юна) будет в следующей статье.
А в данном исследовании авторы, проанализировав генетический профиль императрицы Ашины, пытаются ответить на три ключевых вопроса:
1. Какое было происхождение у древних тюрков?
2. Есть ли генетическая связь между Ашиной и другими тюркскими народами, а также различными популяциями Центральной и Восточной степи после железного века?
3. Есть ли генетическая связь между древними тюрками и современными тюркоязычными народами?
Вступление
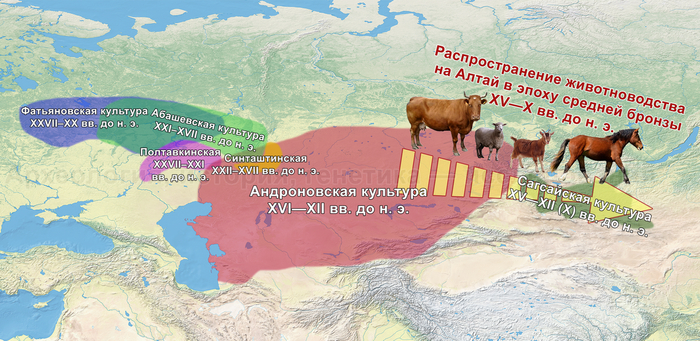
Первый расцвет тюркских народов пришелся на период с VI по VIII век, когда возникла могущественная кочевая империя в виде Тюркского каганата, основанного правителями из рода Ашина, простиравшегося от Монгольского плато до Каспийского моря. Появившись вслед за хунну, сяньби и другими кочевыми конфедерациями, влияние тюрков ощущалось по всей Евразии, что повлияло на языковой ландшафт значительной её части в течение следующих столетий. Тюркский язык, состоящий из более чем 40 языков, является самой многочисленной группой в гипотетической алтайской языковой семье, и около 200 миллионов человек говорят на тюркских языках более чем в 10 странах.
К сожалению, исторические сведения о Тюркском каганате носят разрозненный характер и часто противоречивы. Происхождение древних тюрков является предметом споров, но если не брать мифические истории с тотемными предками по типу появления от хуннского царевича и волчицы, их происхождение может быть сведено к трем конкурирующим гипотезам:
1. Представители племени Ашина выходцы из одноимённого племени хунну в Северо-Восточной Азии.
2. Их происхождение связано с Причерноморско-Каспийской степью или Центральной Азией с последующей миграцией на восток.
3. Сложносоставное происхождение в окрестностях Пинляна или Гаочана в Турфанской впадине, охватывающее как восточные, так и западные евразийские этнические группы.
Поэтому происхождение представителей правящего рода из племени Ашина вызывает большой интерес и является ключом к разгадке происхождения древних тюрков. Тем более с учётом того, что среди тюркской знати была широко распространена кремация.
Результаты
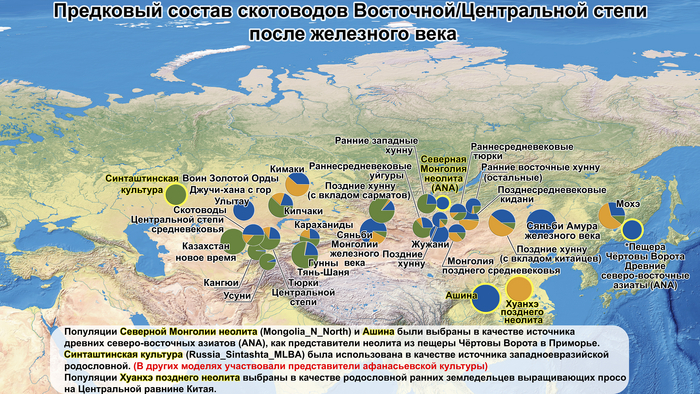
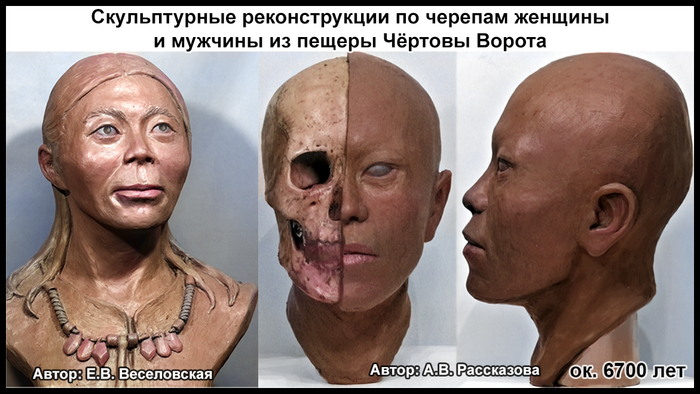
По данным мтДНК императрица Ашина принадлежала к гаплогруппе F1d, распространенной в Северо-Восточной Азии. А по результатам полногеномных анализов она группировалась с древними популяциями Северо-Восточной Азии и востока Монгольского плато, но особенно, с древними северо-восточными азиатами, как представители неолита из пещеры Чёртовы Ворота, возрастом около 6700 лет и носители бойсманской культуры среднего неолита из археологического памятника Бойсмана-2, Приморья, представителями неолита Амура и Северной Монголии, а также с кочевниками Восточной степи после железного века, включая сяньбийцев, жужаней, киданей и части монгольского населения. Из современных популяций она была генетически близка с современными тунгусо-маньчжурскими и монголоязычными народами.
При этом вклада от древних китайцев, связанных с жителями реки Хуанхэ у Ашины не было обнаружено. Однако отмечен небольшой вклад от жителей Западной Евразии – 2,3-3,9%, включающий степную родословную с примесью анатолийских земледельцев, который произошел у предков Ашины за 1566 ± 396 лет до её жизни. В целом генетические данные опровергают гипотезы о западно-евразийском и множественном происхождении представителей знатного рода тюркских каганатов – Ашина в пользу восточноазиатского происхождения.
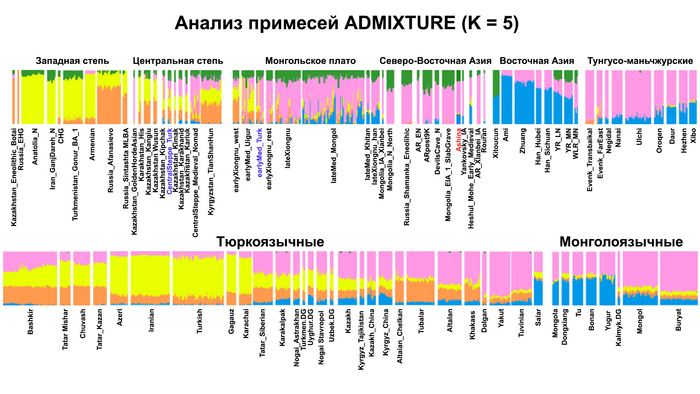
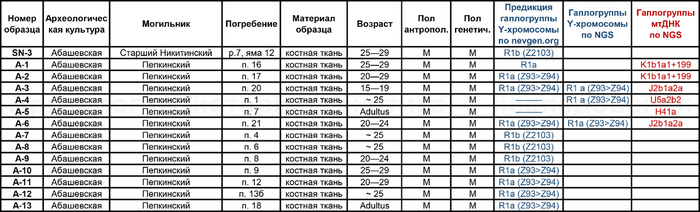
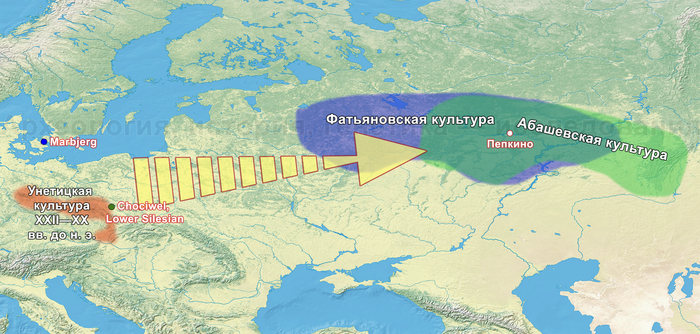
Однако стоит отметить, что у других образцов в тюркский период наблюдалось смешение компонент древних северо-восточных азиатов, которых в среднем было более половины, с компонентами западноевразийских популяций Бактрийско-Маргианского археологического комплекса и кочевников степи железного века, что также соответствует заметному увеличению гаплогрупп Y-хромосомы R и J, в течение тюркского периода в Монголии.
Есть ли генетическая связь между Ашиной и другими тюркскими народами, а также популяциями Центральной и Восточной степи после железного века?
Начиная с железного века в Восточной Степи возникали и рушились, выходящие далеко за её пределы, различные государственные образования кочевников: хунну, сяньби, жужани, тюрки, уйгуры, кидани, кыргызы и – наиболее агрессивные и успешные из всех – монголы. Соседняя средняя часть Евразийской степи или Центральная степь, также была свидетелем перемен в судьбах кочевых образований: гунны, усуни, кангюи, карлуки, кимаки, караханиды и кыпчаки.
Однако представительница элиты Тюркского каганата продемонстрировала генетическую связь только с древнемонгольскими племенами сяньбийцев, киданей и жужаней, а также с представителями тунгусо-маньчжурских племён мохэ. А вот монголы, уйгуры, карлуки, кимаки, кипчаки, а также караханиды были отдалены от кластера Ашины. Хунну вообще был союз разных по происхождению племён, они сильно отличались между собой по географии и по времени. Некоторые западные их представители вообще на 68,4% имели компоненты как у людей из афанасьевской культуры.
Кстати в моделях надо адекватно воспринимать компоненты, они подразумевают схожие генофонды, а не непосредственных представителей культур.
Среди скотоводов Центральной степи усуни, кангюи и гунны Тянь-Шаня большую часть своего происхождения (62,4–73%) ведут от западных степных групп афанасьевской культуры.
А тюркоязычные карлуки, кыпчаки и караханиды, в разных моделях, помимо древних северо-восточных азиатов, имеют 35–50,6% компонент как у афанасьевцев, 10,5–21,7% от жителей Бактрийско-Маргианского археологического комплекса и 38,9–49,4% от выходцев из бассейна реки Хуанхэ железного века. Но доля восточно-евразийского происхождения увеличилась у средневековых скотоводов Центральной степи с 67,3% до 82,5%, за некоторым исключением. Однако стоит отметить, что ещё в новое время на территории Казахстана встречались люди с генетическим профилем на 75,5% происходящем от западных степных скотоводов. При этом Ашина не продемонстрировала близкого генетического родства с тюрками Центральной степи или раннего средневековья. Но в отличие от Ашины, тюрки Центральной Степи и раннесредневековые тюрки продемонстрировали высокую, но вариативную степень западно-евразийского происхождения, что указывает на существование генетической подструктуры ещё у древних тюркоязычных народов.
Есть ли генетическая связь между древними тюрками и современными тюркоязычными народами?
В скотоводческих образованиях Восточной Степи утверждалось, что распространение монгольских языков было связано с сяньбийскими, жужаньскими, киданьскими и монгольскими группами, в то время как хунну, тюрки и уйгуры считались связанными с распространением тюркских языков. В последнем случае были выдвинуты гипотезы о двух волнах распространения: булгаро-тюркской диффузии, начавшейся в гуннский период, спровоцированной более ранней экспансией хунну, и последующей демографической экспансии, связанной с Тюркским каганатом. При этом во втором и третьем веках нашей эры Центральная степь была населена ираноязычными группами, которые постепенно переходили на тюркские языки.
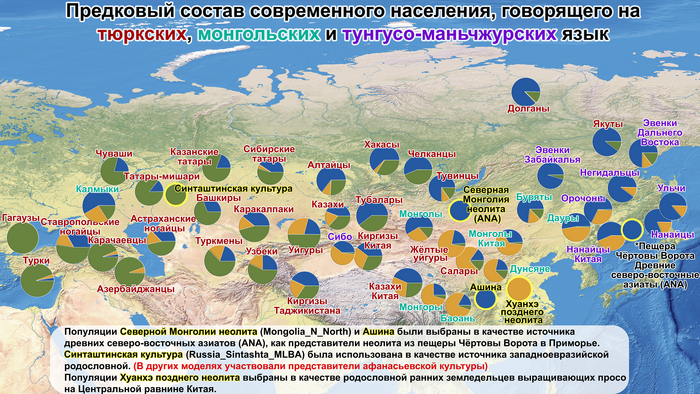
Поэтому генетические данные не подтверждают связи между древними и современными тюркоязычными группами и указывают на значительные различия между тюркоязычными популяциями и популяциями, связанными с распространением тюркских языков.
А пропорции предков древних северо-восточных азиатов, которые составляли основу древних тюрков, варьировались у тюркоязычных групп и даже полностью отсутствовали у некоторых самых западных их представителей. Это указывает на то, что распространение тюркских языков было в основном обусловлено культурными факторами, а не демографической диффузией и интеграцией населения. Поэтому натягивать современные тюркоязычные популяции на тюркизированные древние в качестве доказательства тюркоязычности этих древних популяций абсолютно не имеет смысла и противоречит всем данным.
Пояснение
А подвести итог хочу простым языком, отклонившись от строгой научной линии повествования. Ещё раз: это не научная часть!
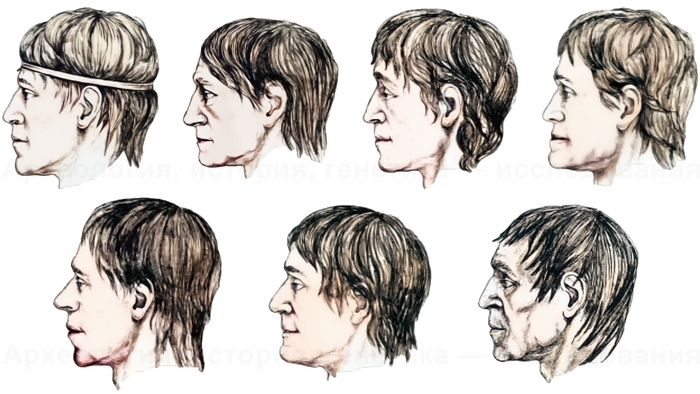
Древние тюрки, и тем более из рода Ашина, по происхождению были близки к древним северо-восточным азиатам, с незначительным вкладом от западных степных скотоводов. По реконструкциям понятно даже как они выглядели. Это характерные отличия.
Древние и современные тюрки имеют разное происхождение. И многие современные тюркоязычные народы объединяет лишь язык.
Отдельные представители тюркских народов в поисках своих предков почему-то обращаются к древним тюркам, однако схожих с ними компонент у них не так много, а у некоторых вообще почти нет. Их генофонд состоит из смеси различных древних популяций, говоривших на разных языках, при этом у разных народов и разный генетический коктейль и своя интересная история. Но если древних тюрков начинать искать от современных, то можно выйти на разные популяции, которые внесли свой вклад в современные и намеренно или искренне заблуждаясь, объявить их тюрками.
Характерным примером таких манипуляций выглядит якобы доказательство тюркоязычности скифов, основанное только на словах из исследования 2017 года, что современные потомки западно-скифских групп встречаются среди различных современных групп на Кавказе и в Центральной Азии, в то время как сходство с восточно-скифскими группами встречается более широко, но почти исключительно среди современных тюркоязычных групп, особенно из кыпчакской ветви тюркского языка, которые в прошлом были кочевниками. Что не удивительно с учётом огромного генетического разнообразия этих групп, значительно большего чем у всех народов Европы вместе взятых. Трудно промахнуться в стадо слонов из дробовика. К тому же ещё и с учётом того, что сами тюркоязычные карлуки, кыпчаки и караханиды, в разных моделях, помимо древних северо-восточных азиатов, как уже было сказано выше, имеют 35–50,6% предков от афанасьевцев, 10,5–21,7% от жителей Бактрийско-Маргианского археологического комплекса и 38,9–49,4% от выходцев из бассейна реки Хуанхэ железного века. Но они же по определению тюрки, и мы не называем кыпчаков индоевропейцами или монголами с китайцами! С такой логикой если в Анадыре с вами поздоровался и разговаривает чукча на чисто русском языке, то его предки по мнению таких вот специалистов в кавычках, должны быть исключительно русскими.
Работа о скифах 2017 года: Unterländer, M., Palstra, F., Lazaridis, I. et al. Ancestry and demography and descendants of Iron Age nomads of the Eurasian Steppe. Nat Commun 8, 14615 (2017). https://doi.org/10.1038/ncomms14615
И это исследование 2017 года в комментариях, не цитировал разве что только ленивый защитник тюркского происхождения скифов и не только. И продолжат это делать, вот увидите. При этом само исследование они не читали и вряд ли понимают…
Толкование скифских слов из тюркского несостоятельно хотя бы потому, что фонетика засвидетельствованных скифских слов противоречит фонетическим характеристикам пратюркского (распад которого по глоттохронологическим данным относится к I в. до н. э., т. е. много позже появления на исторической арене скифов – соответственно, сравнивать скифские слова имело бы смысл только с пратюркскими, а не с их рефлексами в отдельных древних и современных тюркских языках). Так, пратюркское слово не могло начинаться на сонанты *m
и *l, тогда как нам известны скифские имена Мадий и Липоксай; невозможными в пратюркском были и скопления согласных в анлауте (начале слова), засвидетельствованные в самоназвании скифов и таких именах, как, например, Спаргапиф. Одним словом, попытки объявить скифов тюрками представляют интерес разве что с точки зрения современной этнопсихологии. Сергей Кулланда http://генофонд.рф/?page_id=3267
1/3
Интервью одного из авторов работы о скифах 2017 г., руководителя межинститутского сектора молекулярной палеогенетики Института цитологии и генетики СО РАН, Института археологии и этнографии СО РАН Александра Пилипенко на научно-популярном сайте elementy.ru
К примеру, от союза русского и китаянки положение их детей может быть на том же графике где-то около ногайцев или ближе к казахам.
*Очень грубый пример
И по пропорциям главных компонент они будут на них похожи, но ведь нельзя же будет их ребенка назвать казахом или ногайцем. Часто вижу непонимание этого элементарного биологического аспекта в комментариях. Но когда нужно что-то доказать о себе любимом и, тем более, великом, тут логика ломается.
Или упор часто делают на то, что если кто-то до наших дней сохранил традиции, связанные с кочевым образом жизни, питанием и тому подобно… То только его предки в прошлом могли этим заниматься. Мне вот интересно, чем занимались по такой логике предки тех, кто сейчас компьютерные технологии развивает или в космос ракеты запускает? А так и бомжам можно сказать, что они продолжатели славных традиций охотников и собирателей… Уж пусть не обижаются на меня современные кочевники и скотоводы, а также их потомки. В сохранении традиций нет ничего плохого, если не сочинять себе историю…
Примечание: текста много, тема сложная и состоит из двух постов. Разделы – "Социокультурные изменения" и "Итоги" (Появление и сокращение генетического разрыва), во второй части.
Вступление
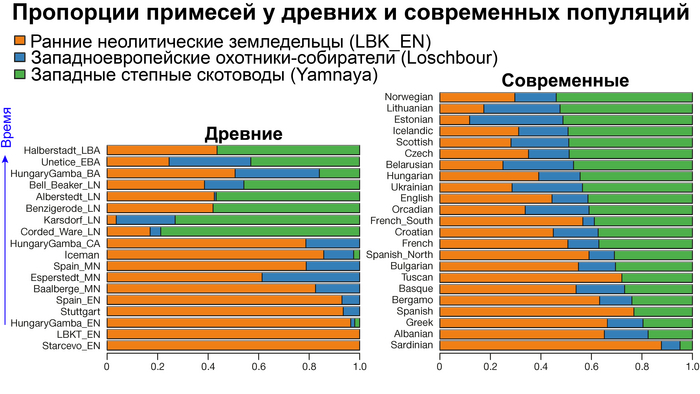
Генетическое разнообразие человеческих популяций Западной Евразии в значительной степени сформировалось в результате трех основных доисторических миграций:
1. Миграции охотников-собирателей, а именно анатомически современных людей, находящихся на ранней стадии социально-экономического развития, которые занимали территорию, примерно, с 45 тыс. лет назад. При этом вклад в последующие популяции внесли выжившие их линии после последнего ледникового максимума около 25-19 тыс. лет назад.
2. Далее, гораздо ещё более существенный вклад, внесли ранние неолитические земледельцы, пришедшие с Ближнего Востока примерно с 11 тыс. лет назад.
3. Степные скотоводы, распространившиеся из Причерноморско-Каспийской степи около 5000 лет назад, привнесли третью основную генетическую компоненту в популяции Западной Евразии.
Аутосомный генофонд практически всех европейских популяций по большей части состоит из различных пропорций этих предковых компонент, при этом независимо какие у них гаплогруппы.
Европейский континент заселялся людьми современной анатомии в несколько волн, при этом не все линии этих людей были предками современных. Первые «сапиенсы» Европы (более 40 тыс. лет назад) пересекались и скрещивались с неандертальцами, пока те совсем не вымерли. Об этой примеси свидетельствуют геномы людей из пещеры Бачо Киро в Болгарии, связанных с археологической культурой начального верхнего палеолита, и со стоянки Пештера-ку-Оасе в Румынии. Другие люди того периода, такие как женщина из пещеры Златый кунь Чехии и человек из Усть-Ишима России, не несут в своих геномах значительного количества неандертальского вклада, чем другие неафриканские группы. При этом люди, жившие в Европе более 40 тыс. лет назад, не оставили существенных следов в генетическом составе современных евразийских популяций, что говорит о том, что они в значительной степени вымерли или были ассимилированы последующими группами людей современной анатомии.
В те времена, на демографию древних людей влияли различные факторы, в том числе и климатические. Пик похолодания во время последнего ледникового максимума (LGM) был около 21 тыс. лет назад, когда Северная Европа была в значительной степени покрыта льдом, а южная граница ледниковых щитов проходила через Германию и Польшу. Многие участки Европы стали холодными пустынями с тундростепями и лесостепями на побережье Средиземного моря. А люди и животные переместились в южные широты, о чем свидетельствует, например, солютрейская культура на Пиренейском полуострове и на юге Франции. Другими предложенными рефугиумами или климатическими убежищами в тот период были Апеннинский полуостров, Балканы и юго-восточная часть Европейской равнины.
А в результате послеледниковой колонизации, палеогенетический анализ выявил разделение популяций охотников-собирателей на западных и восточных. Источники родословной западных охотников-собирателей (WHG) связаны с носителями эпиграветтской, азилийской и других эпипалеолитических культур генетического кластера Виллабруна, появление которого обусловлено миграциями в конце ледникового периода и появлением новой генетической компоненты, связанной с Ближним Востоком того времени, начиная с бёллинг-аллерёдского потепления около 14 тыс. лет назад, что археологически коррелирует с культурными переходами.
А родословная восточных охотников-собирателей (EHG) демонстрирует дальнейшую примесь с верхнепалеолитическими популяциями Сибири - древними северными евразийцами (ANE).
Предковый состав западных охотников-собирателей (WHG) был регионально изменчивым в мезолитических популяциях. Если охотники и собиратели из Британии и северо-западной континентальной Европы продемонстрировали однородный состав от палеолита и мезолита, а также вплоть до неолитической экспансии около 6000 лет назад, то их коллеги из Испании, представляли собой смесь людей из двух рефугиумов, когда к генетическому профилю кластера Виллабруна, добавляется примесь от более ранних местных пиренейских охотников-собирателей, связанных с солютрейской и мадленской культурами верхнего палеолита.
Этот генетический профиль сохранился во время последнего ледникового максимума в Юго-Западной Европе около 25-19 тыс. лет назад.
А время появления примеси сформировавшей восточных охотников-собирателей (EHG), оценивается в 15–13 тыс. лет назад, однако её пропорции в популяциях, по-видимому, соответствуют клине, которая в целом коррелирует с географией, так охотники-собиратели Прибалтики и Украины демонстрируют большее сходство с предками кластера Виллабруна, чем охотники и собиратели западной части современной России, у которых было больше вклада от древних северных евразийцев (ANE).
В свою очередь скандинавские охотники и собиратели продемонстрировали различные пропорции смеси западных и восточных охотников и собирателей (WHG и EHG) в позднем мезолите.
Стоит отметить, что охотники и собиратели мезолита и тем более палеолита, были очень разнообразны и неоднородны, но из-за ограниченного количества данных, наши знания о мезолитической структуре населения и процессах демографического смешения долгое время были скудны и содержали существенные хронологические и географические пробелы в информации. Отчасти это объясняется малым количеством хорошо сохранившихся мезолитических человеческих останков возрастом более 8000 лет, а отчасти и потому, что большинство исследований древней ДНК, посвященных периодам мезолита и неолита, были ограничены преимущественно находками из Европы.
1/3
Хотя давно напрашивается более детальная классификация охотников и собирателей по их характерным отличиям. Так, археологические данные указывают на условную границу от восточной Балтики до Черного моря, к востоку от которой сообщества охотников и собирателей существовали гораздо дольше, чем в Западной Европе, несмотря на аналогичное расстояние до центра распространения раннего земледелия на Ближнем Востоке.
Компоненты восточного и западного происхождения у охотников-собирателей в этом пограничном регионе сильно различаются, но более точные представления о демографических процессах, связанных с разделением на восток и запад и о динамике численности населения к востоку от Европы, включая Северную и Центральную Азию за тот же период времени, весьма ограничены. В этих регионах термин «неолит» характеризуется культурными, экономическими и социальными изменениями, а также использованием керамики. К примеру, неолитические культуры степей Центральной Азии и таежного пояса России уже изготавливали керамику, однако продолжали вести прежний образ жизни с присваивающей экономикой, как у предшествующих мезолитических культур охотников-собирателей, с различными инновациями в зависимости от условий среды обитания. Однако пока недостаточно данных для понимания того, как неолитизация отличалась по своим срокам, механизмам и последствиям в разных уголках Евразии.
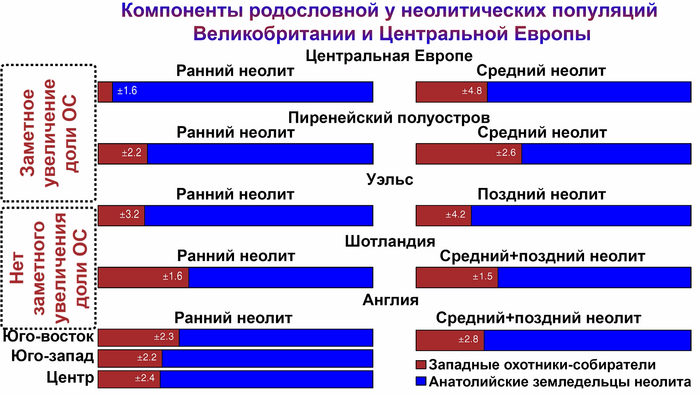
В Европе, кроме восточной её части, переход от охоты и собирательства к земледелию сопровождался прибытием мигрантов ближневосточного происхождения, практикующим производящее хозяйство на основе одомашненных растений и животных. С последующим значительным ростом населения. В Юго-Восточной, Центральной и Западной Европе анатолийские земледельцы впоследствии в некоторой степени смешались с местными охотниками и собирателями, а на территории Британии, прибывшие с континентальной Европы ранние европейские земледельцы полностью заменили местных охотников и собирателей.
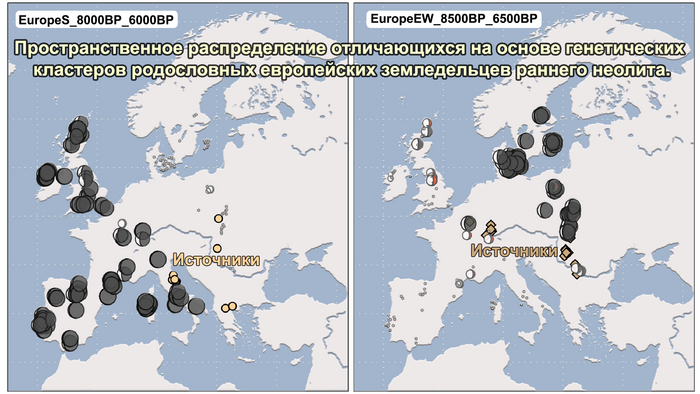
А вот в восточно-балтийском регионе наблюдается заметно иная модель неолитизации, с появлением одомашненных животных только при появлении культуры шнуровой керамики (CWC) около 4800 лет назад. Аналогичным образом в Северном Причерноморье люди с генетическим профилем мезолитических охотников и собирателей тысячелетиями сосуществовали с земледельческими группами, расположенными дальше на запад.
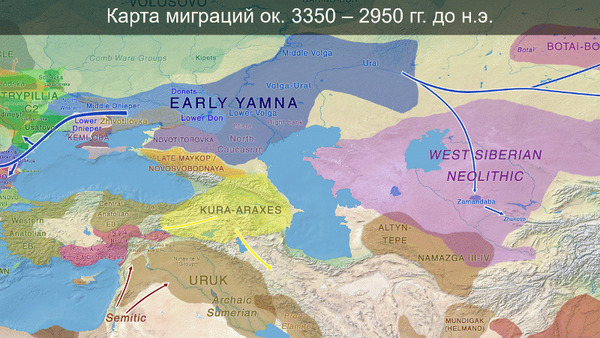
И третий крупнейший генетический пласт в истории европейских популяций связан со степными скотоводами раннего бронзового века, как представители ямной культуры около 5 тыс. лет назад. Этот генетический профиль, быстро распространился по Европе посредством экспансии как носителей культуры шнуровой керамики (CWC), так и родственных ей культур.
Хотя предыдущие исследования выявили эти крупномасштабные миграции в Европу и Центральную Азию, основные аспекты, касающиеся демографических процессов, не решены. Ямная или степная родословная в общих чертах охарактеризована как смесь восточноевропейских (EHG) и кавказских охотников-собирателей (CHG), образовавшаяся в результате гипотетической смеси «северного» степного и «южного» кавказского источников. Однако точное происхождение этих предковых источников не было установлено. Более того, за некоторым исключением, известные гаплогруппы Y-хромосомы представителей ямной культуры не соответствуют гаплогруппам, обнаруженным у европейцев после 5000 тыс. лет назад и происхождение этой отцовской линии также не установлено. К тому же "степная" родословная в Европе до сих пор была выявлена только в смешанной форме, а детали и механизмы, с помощью которых эта родословная впоследствии распространилась вместе культурой шнуровой керамики (CWC), оставались неясными.
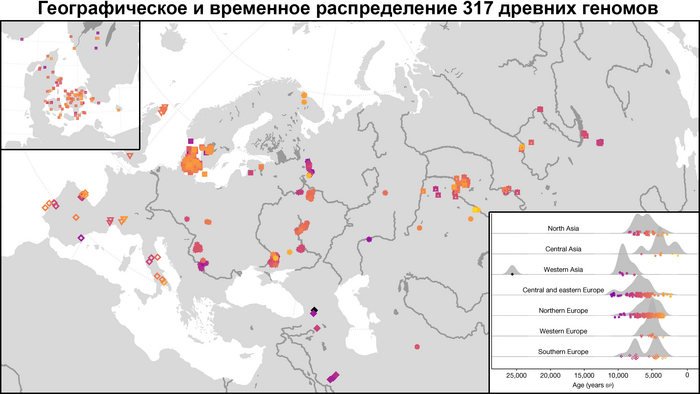
В новой работе учёные под руководством Эске Виллерслева из образцов, датированных радиоуглеродным методом при помощи ускорительной масс-спектрометрии, секвенировали геномы 317 человек, 309 из них датируются периодом от 11 до 3 тысяч лет назад.
А совместно с ранее опубликованными данными получилась выборка из 1600 древних геномов, секвенированных с помощью метода дробовика.
Географически 317 отобранных образцов охватывают обширную территорию Евразии, по долготе от Атлантического океана до озера Байкал, а по широте от Скандинавии до Ближнего Востока, и происходят из контекстов, которые включают курганы, пещеры, болота и дно водоёмов.
Географически образцы можно разделить на три региона:
1. Центральная, Западная и Северная Европа.
2. Восточная Европа, включая западную часть России, Беларусь и Украину.
3. Урал и Западная Сибирь.
Образцы включают представителей многих ключевых культур Западной Евразии мезолита, неолита и последующих периодов, такие как маглемозе, эртебёлле, воронковидных кубков (TRB) и шнуровой керамики или одиночных погребений в Скандинавии; импрессо или кардиумной керамики в Средиземноморье; кёрёш и линейно-ленточной керамики (LBK) в Юго-Восточной и Центральной Европе; и многие археологические культуры Украины, западной части России и Зауралья (как веретьё, льяловская, волосовская, китойская и др.). Выборка была особенно плотной в Дании, где сотня геномов, охватывала период от раннего мезолита до бронзового века. Обширная выборка была также получена из Украины, Западной России и Зауралья, охватывая период от раннего мезолита до неолита, примерно до 5000 лет назад.
Результаты
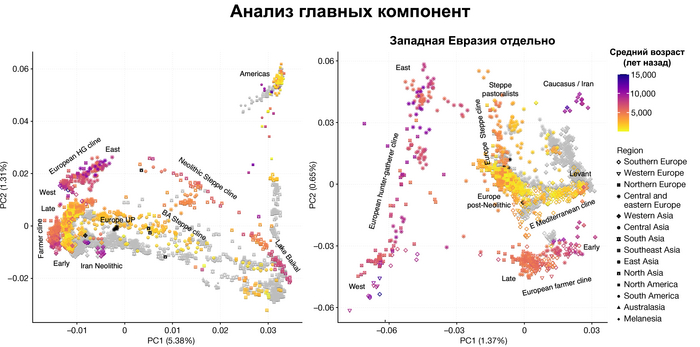
Полученный набор генетических данных позволил при анализе главных компонент использовать за основу древние геномы, и уже на них спроецировать положение современных популяций, а не наоборот, как это было во многих предыдущих исследованиях.
И хотя результаты согласуются с тем, что древние люди сильнее друг от друга генетически отличались, чем современные, такой подход позволил выявить ещё более значительные генетические различия между древними людьми, которые ранее не наблюдались, что отражает меньший эффективный размер их популяции и большую генетическую изоляцию.
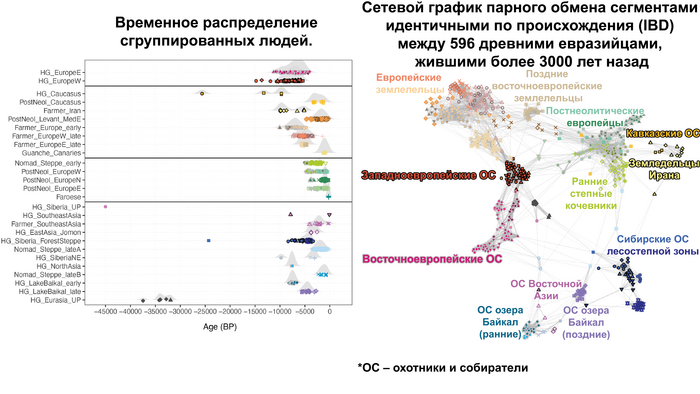
Чтобы получить более точную генетическую характеристику в пространстве и времени, авторы использовали попарную идентификацию сегментов ДНК идентичных по происхождению, чтобы сгруппировать анализируемых людей в связанные кластеры сходного генетического происхождения, а не по культурным или территориальным признакам.
В более крупном масштабе они отражали ранее описанные группы западных и восточных охотников-собирателей, а вот в более мелком масштабе они сгруппировались не только в пределах ограниченных пространственно-временных диапазонов и/или археологических контекстов, но также продемонстрировали ранее неизвестные связи в более широких географических диапазонах.
Полученные кластеры далее использовались для моделирования состава предков последующих популяций.
Популяционная структура охотников и собирателей после максимума последнего оледенения (LGM)
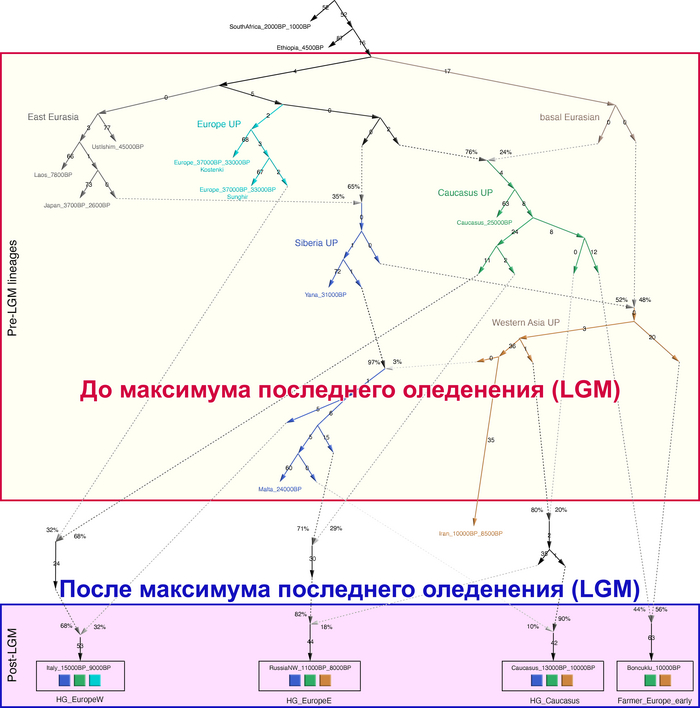
До последнего ледникового максимума, популяции Евразии довольно сильно отличались и в разной степени смешивались с неандертальцами. Некоторые популяции пока остаются гипотетическими, как к примеру, так называемые, «базальные евразийцы» с незначительной или нулевой примесью от неандертальцев, их прямых образцов, подтвержденных генетикой, на данный момент нет, но генетические следы этой популяции были обнаружены у охотников-собирателей Кавказа и Западной Азии верхнего палеолита, а также у более поздних западноевропейских охотников и собирателей.
В данной работе авторы сообщают о генетических данных полученных из зуба женщины верхнего палеолита, жившей около 26-25 тыс. лет назад в окрестностях карстового грота Котиас Клде в современной Грузии, не путать с более молодыми образцами кавказских охотников-собирателей из этого археологического комплекса, у которых было менее 20% этой кавказской родословной верхнего палеолита. А сам геном этой представительницы Кавказа верхнего палеолита представляет собой смесь 76% западноевразийских охотников-собирателей верхнего палеолита (т.е. до максимума последнего оледенения) и 24% «базальных евразийцев». Кстати популяции Ближнего Востока верхнего палеолита, почти наполовину произошли от «базальных евразийцев», которые, практически не имели примеси неандертальцев и отделились от других неафриканских линий ещё до их разделения между собой. При этом западноевропейские охотники-собиратели, жившие после последнего ледникового максимума, моделируются как итоговая, за несколько событий, смесь 46% популяций, связанных с Кавказом верхнего палеолита, 22% европейцев, живших около 33-37 тыс. лет назад, связанных со стоянками Сунгирь и Костёнки и 32% древних популяций, связанных с Сибирью верхнего палеолита, как представитель стоянки Мальта, возрастом 24 тыс. лет, т.е. древних северных евразийцев.
В свою очередь восточноевропейские охотники-собиратели моделируются как смесь 58% популяций, связанных с обитателями стоянки Мальта в Прибайкалье, 28% популяций, связанных с Кавказом верхнего палеолита, как женщина из Котиас Клде (но в разные периоды, сначала 24%, а потом 4%) и 14% представителей Западной Азии верхнего палеолита. Эта смесь также проходила в несколько этапов. Тем самым новые данные подтверждают генетическое разделение охотников и собирателей в Восточной и Западной Европе после максимума последнего оледенения, когда люди снова расселялись по Евразии, при этом эти различия сохранялись на протяжении всего мезолита.
В анализе примесей, эти сложносоставные генофонды представлены отдельными компонентами. А различия между ними, а именно между западноевропейскими и восточноевропейскими охотниками-собирателями мезолита максимально выражены у представителей Италии, возрастом от 15 до 9 тыс. лет и Северо-Запада современной России возрастом от 11 до 8 тыс. лет.
Обратите внимание, что генетический состав одинаков даже у людей с разными гаплогруппами и наоборот, по всему геному люди с одинаковыми линиями гаплогрупп отличаются.
Кстати, в представлении сложных процессов, не стоит забывать о временных промежутках, о которых идёт речь.
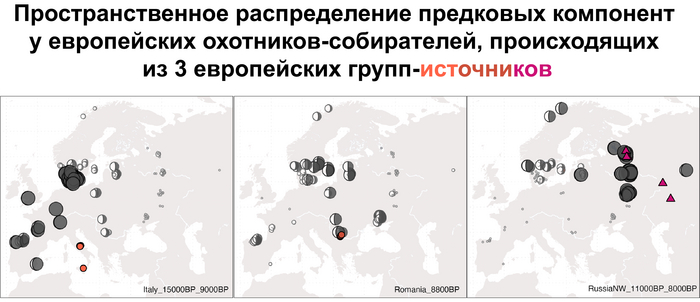
Что касается североевропейских охотников и собирателей, то они отличались по пропорциям смеси компонент как у западноевропейских и восточноевропейских охотников-собирателей, за исключением Дании, где популяции продемонстрировали южноевропейское происхождение и были генетически схожи с охотниками-собирателями из Италии, возрастом от 15 до 9 тыс. лет. При чём эта родословная сохранялась в регионе на протяжении 5 тыс. лет.
В целом результаты указывают на то, что существовало по меньшей мере три отчетливые волны миграций охотников и собирателей в Скандинавию:
1. Первая волна связана с миграцией людей южноевропейского происхождения на территорию Дании и прибрежной части Юго-Западной Швеции.
2. Вторая волна в Прибалтику и Юго-восточную Швецию связана с популяциями из Юго-Восточной Европы.
И 3. Третья волна миграций на крайний север Скандинавии, а затем на юг вдоль атлантического побережья Норвегии связана с охотниками и собирателями с северо-запада современной России.
Эти миграции, вероятно, представляют собой послеледниковую экспансию из рефугиумов или районов-убежищ, общих как для людей, так и для многих видов растений и животных.
Стоит отметить, что генофонд европейских охотников-собирателей верхнего палеолита, живших до максимума последнего оледенения был в значительной степени заменён генофондом, происходящим из рефугиума в Южной Италии. На Пиренейском полуострове самые древние образцы из этого исследования, в том числе охотника-собирателя, возрастом около 9500 лет (NEO694) из бухты Санта-Майра на востоке Испании, показали преимущественно южноевропейское происхождение с незначительным вкладом охотников и собирателей верхнего палеолита, который, вероятно, отражает генофонд, связанный с мадленским периодом до максимума последнего оледенения.
А более поздние жители севера Пиренейского полуострова, были ближе к охотникам и собирателям Юго-Восточной Европы, демонстрируя около 40–30% вклада балканских охотников-собирателей.
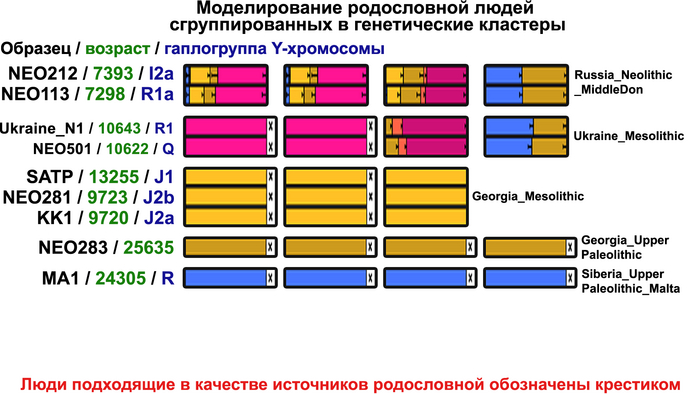
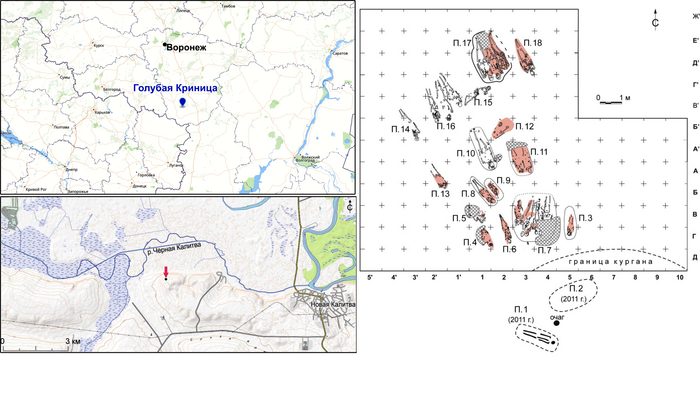
Поток генов, связанных с охотниками-собирателям Юго-Восточной Европы был направлен и в Восточную Европу после мезолита, на территорию современной Украины и далее. А у двух человек из могильника мариупольского типа раннего энеолита (NEO212, NEO113 ) близ хутора Голубая Криница на юге Воронежской области, возрастом около 7300 лет, генетический профиль состоял уже из смеси более ранних охотников-собирателей с территории Украины с вкладом от 18 до 24% из источника, связанного с охотниками-собирателями Кавказа.
1/2
Поэтому на графике анализа главных компонент они отклонены в сторону Ирана и Кавказа. По расчётам эта примесь произошла примерно 8300 лет назад. Результаты указывают на более ранний генетический контакт между популяциями Кавказа и степи ещё до появления культур кочевников в Причерноморско-Каспийской степи. Помимо этого, этот контакт задокументирован западнее, чем предполагалось ранее.
Основные генетические изменения в Европе
Предыдущие исследования древней геномики задокументировали несколько эпизодов крупномасштабной смены населения в Европе за последние 10 тыс. лет, но 317 новых геномов из данной работы, заполняют важные пробелы в наших знаниях.
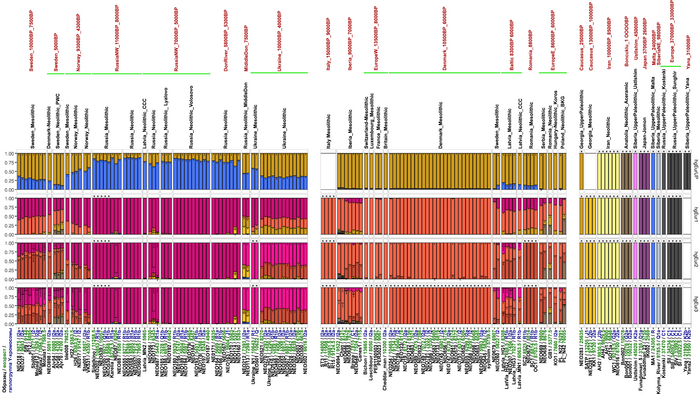
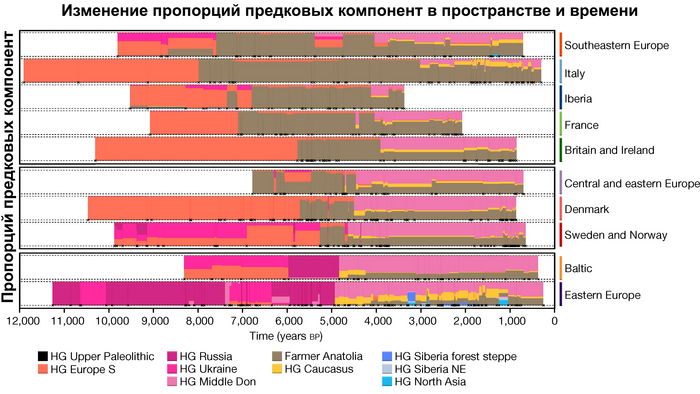
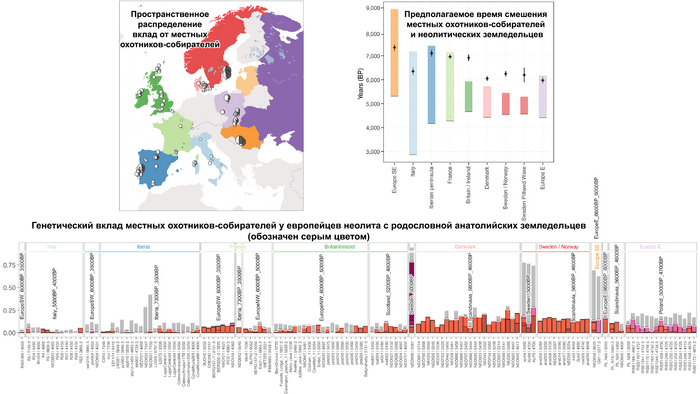
Анализ выявил глубокие различия в пространственно-временной динамике неолитизации между восточной и западной частями Европы вдоль пограничной зоны, простирающейся от Черного моря до Балтийского. К западу от этой пограничной зоны, переход к неолиту сопровождался крупномасштабными сдвигами в генетической родословной, от местных охотников-собирателей к земледельцам анатолийского происхождения (Boncuklu_10000BP). Этот переход в зависимости от региона охватывал обширный период времени, более 3 тыс. лет, от самых ранних свидетельств на Балканах около 8700 лет назад до 5900 лет назад в Дании.
В последующие периоды смешение между ранними европейскими земледельцами и местными охотниками-собирателями привело к частичному увеличению родословной охотников и собирателей во многих регионах Европы, но этот вклад редко превышал 10%, помимо заметных исключений, наблюдаемых у людей из района Железных ворот на границе современных Сербии и Румынии, а также у носителей культур ямочной керамики из Швеции и кардиумной керамики раннего неолита из Португалии, которые, по оценкам, содержат от 27 до 43% вклада от охотников и собирателей, как представители Пиренейского полуострова, возрастом от 9 до 7 тыс. лет (Iberia_9000BP_7000BP).
Обитатели эпохи неолита с территории современной Дании продемонстрировали до 25% генофонда охотников-собирателей (EuropeW_13500BP_8000BP), но преимущественно не местных.
Также обнаружены региональные различия в происхождении последующих неолитических групп. Так последующие неолитические популяции Западной Европы происходят от ранних земледельцев Южной Европы. А в последующие неолитические группы Восточной Европы и Скандинавии внесли свой вклад преимущественно ранние земледельцы Центральной Европы.
А вот с восточной стороны упомянутой пограничной зоны, на территории Прибалтики, Украины и западной части России, не наблюдается существенных генетических изменений, а предковые линии местных охотников и собирателей, без заметного вклада от анатолийских земледельцев,сохраняются примерно до 5 тыс. лет назад. Эта генетическая преемственность согласуется с археологическими данными, которые свидетельствуют о сохранении в этом обширном регионе групп охотников, рыболовов и собирателей, которые использовали керамику, но не практиковали земледелие и животноводство ещё на несколько тысяч лет.
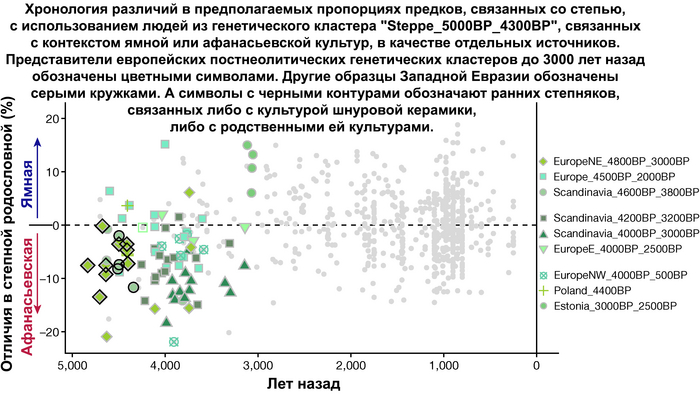
Но несмотря на то, что влияние степной родословной на популяции Евразии хорошо известно, вопросы о её происхождении оставались не решенными. В данной работе авторы показывают, что состав степной родословной (Steppe_5000BP_4300BP) может быть смоделирован как смесь примерно 65% компонент, связанных с охотниками-собирателями, использующими керамику, из района Среднего Дона, как люди из могильника мариупольского типа Голубая Криница раннего энеолита на юге Воронежской области, возрастом около 7300 лет (MiddleDon_7500BP) и 35% компонент, связанных с охотниками и собирателями Кавказа, возрастом от 13 до 10 тыс. лет (Caucasus_13000BP_10000BP).
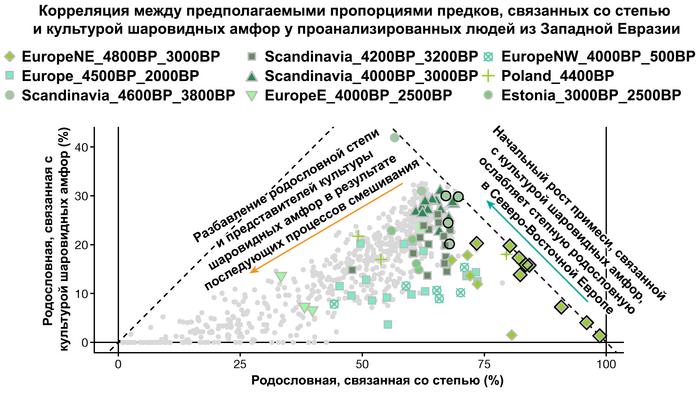
Таким образом, охотники-собиратели Среднего Дона, которые уже имели родословную, связанную с кавказскими охотниками-собирателями, являются неизвестным до сих пор ближайшим так называемым северным источником большей части родословной у людей, связанных с ямной культурой. При этом генетический состав людей из могильника Голубая Криница, отличается от такового у других древних людей из могильников мариупольского типа Приднепровья. Однако распространение степной родословной в Европе связно с культурой шнуровой керамики, носители которой представляют собой смесь предковых линий, связанных со степью и неолитическими земледельцами, происхождение которых может быть связано исключительно с культурой шаровидных амфор.
Это наблюдение имеет большое значение для понимания возникновения культуры шнуровой керамики. Хотя уже давно высказывались предположения о стилистической связи между керамикой культур шаровидных амфор и шнуровой керамики, включая использование сосудов в форме амфор и узоры, выполненные шнуром. При этом незадолго до появления самых ранних групп культуры шнуровой керамики восточные группы культуры шаровидных амфор и западные группы ямной культуры обменялись культурными элементами в лесостепной переходной зоне к северо-западу от Черного моря, где керамические амфоры и кремневые топоры культуры шаровидных амфор были включены в погребения ямной, а типичное для ямной использование охры встречалось в погребениях культуры шаровидных амфор, что указывает на тесное взаимодействие между этими группами. Предыдущие генетические анализы нескольких людей из зоны контакта предполагали, что эти контакты были ограничены культурным влиянием, а не примесью. Однако, в свете новых генетических данных, похоже, что эта зона — и, возможно, другие подобные зоны контакта между культурой шаровидных амфор и группами из степи (как из ямной культуры) — были ключевыми в формировании культуры шнуровой керамики, благодаря которой генетические компоненты, связанные со степью, и культурой шаровидных амфор, распространились далеко на запад и север, что привело к регионально разнообразным ситуациям взаимодействия и смешения.
При этом распространение культуры шнуровой керамики происходило преимущественно благодаря связям, которые были выстроены культурой шаровидных амфор в предшествующий период. Но несмотря на сходство по аутосомным данным, различные линии гаплогрупп Y-хромосомы у представителей культур шнуровой керамики и ямной предполагают, что известные на данный момент образцы представителей ямной культуры не подходят в качестве непосредственного источника степной родословной у носителей культуры шнуровой керамики.
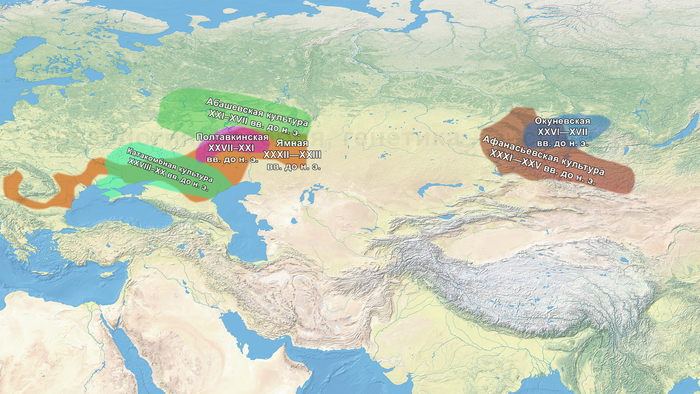
Это предположение было подтверждено моделированием предковых компонент у европейцев со степным происхождением возрастом от 3 тыс. лет назад с участием представителей родственных ямной и афанасьевской культур, потому как модели показывают, что представители афанасьевской культуры, хоть и незначительно, но оказались более лучшим источником степной родословной у европейцев, несмотря на разное географическое распространение.
Стабильность популяций охотников-собирателей к востоку от Урала
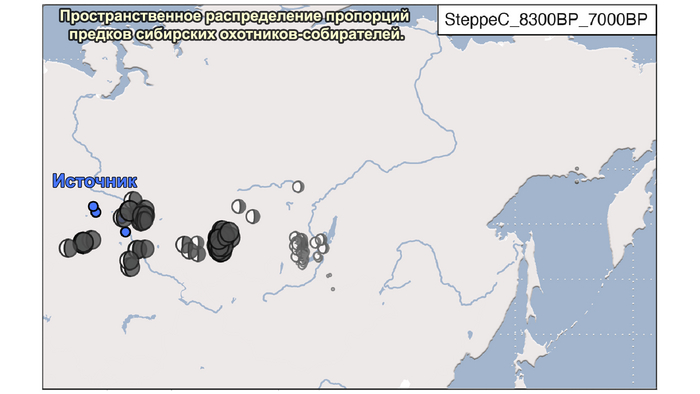
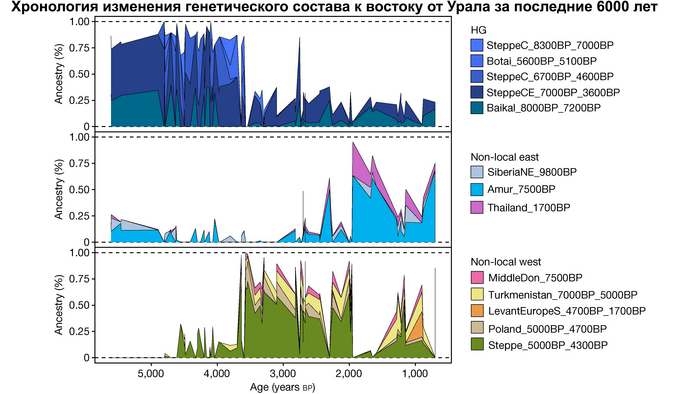
В отличие от значительного числа геномов древних охотников-собирателей из Западной Евразии, которые были изучены до настоящего времени, геномные данные охотников-собирателей к востоку от Урала были скудны. Эти регионы характеризуются ранним появлением керамики из районов, расположенных дальше к востоку, они были населены сложными обществами охотников-собирателей с постоянными, а иногда и укрепленными поселениями. В данной работе сообщается о 38 новых образцах, 28 из которых от охотников-собирателей, использующих керамику 8300-5000 лет назад. Большинство этих геномов попадает на неолитическую степную клину, которая охватывает сибирские лесостепные зоны бассейнов рек Иртыш, Ишим, Обь, Енисей, а также озера Байкал.
Эти охотники и собиратели могут быть смоделированы как смесь компонент из трёх генетических кластеров:
1. Компонента западносибирских охотников-собирателей возрастом от 8300 до 7000 лет (SteppeC_8300BP_7000BP), доминировала в западной части лесостепной зоны.
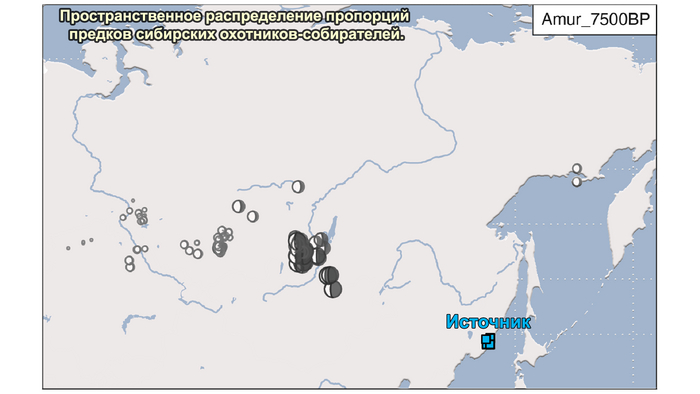
2. Компонента охотников-собирателей Северо-Восточной Азии возрастом около 7500 лет (Amur_7500BP) или древних северо-восточных азиатов из пещеры Чёртовы ворота в Приморье, была максимально выражена у популяций Байкала.
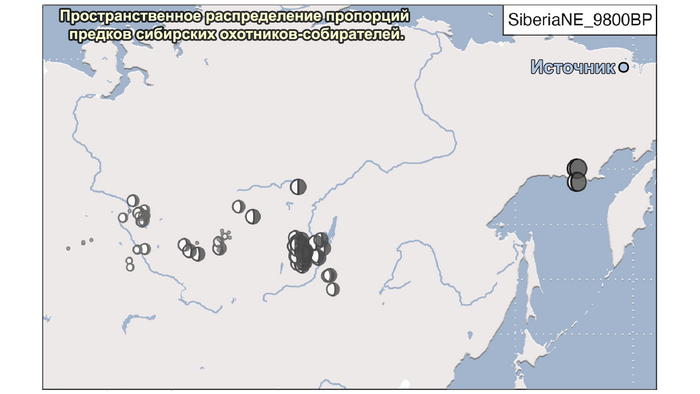
И 3. Компонента представлена палеосибирской родословной возрастом 9800 лет (Siberian_9800BP) как у обитателя Дуванного Яра в низовьях реки Колымы, она снижались по мере удаления от озера Байкал на запад.
1/5
Авторы использовали кластеры также и для исследования пространственно-временной динамики генетического состава в степи и районе озера Байкал после неолита. Результаты подтверждают увеличение компоненты охотников-собирателей лесостепи возрастом от 7000 до 3600 лет (SteppeCE_7000BP_3600BP) у жителей Байкала позднего неолит-ранней бронзы (кластеры Baikal_5600BP_5400BP и Baikal_4800BP_4200BP). Однако родословная, относящаяся к этому кластеру, уже наблюдалась у вновь описанных неолитических охотников-собирателей как у озера Байкал (NEO199 и NEO200), так и вдоль Ангары на севере (NEO843) около 7000 лет назад. У двух мужчин Байкала была определена гаплогруппа Y-хромосомы Q1b1, характерная для последующих групп позднего неолита-ранней бронзы из того же региона. Что совместно с расчётной датой примеси около 7300 лет назад, позволяет предположить, что поток генов между охотниками-собирателями Байкала и лесостепей Южной Сибири уже был во времена раннего неолита, что согласуется и с археологическими данными.
Новые образцы также проливают свет на генетическое происхождение носителей окуневской культуры раннего бронзового века в Минусинской котловине на юге Сибири. Её представителей можно смоделировать как смесь сибирских охотников-собирателей лесостепной зоны из кластеров возрастом 6700-4600 и 7000-3600 лет (Steppe_6700BP_4600BP и SteppeCE_7000BP_3600BP) со степными скотоводами возрастом от 5000 до 4300 лет (Steppe_5000BP_4300BP), связанными исключительно с афанасьевской культурой, время этой примеси датировано около 4600 лет назад.
1/2
А примерно 3700 лет назад жители в степном регионе и у озера Байкал демонстрируют уже заметно отличающиеся генетические профили. Ранние стадии этого перехода характеризуются притоком предков, связанных со степью, количество которых снижается со временем, от 70% на начальном этапе.
Аналогично динамике в Западной Евразии, происхождение, связанное со степью, здесь соотносится с таковым как у представителей культуры шаровидных амфор с территории современной Польши, возрастом 5000-4700 лет, подтверждая ранее задокументированный поток генов от групп из культуры шаровидных амфор в соседние группы степи и лесостепи, а также поток этой уже смешанной с европейскими земледельцами родословной дальше на восток уже с синташтинской и андроновской культурами в бронзовом веке. Однако у людей из окуневской культуры отсутствует вклад от людей культуры шаровидных амфор, а люди со степным происхождением начиная от 3700 лет назад демонстрируют чуть большую близость к ямной, чем к афанасьевской культуре, что указывает на две волны миграций западных степных скотоводов на восток. Первая волна в раннем бронзовом веке была связана с ямной культурой, а вторая, позже, была связана с синташтинской и андроновской культурами. Более поздние этапы переходного периода характеризуются увеличением родословной, связанной с иранскими земледельцами Центральной Азией (Turkmenistan_7000BP_5000BP) и древними северо-восточными азиатами Приморья (Amur_7500BP).
Суммарно, эти результаты показывают, что на востоке Евразийской степи родословная охотников и собирателей со сложной генетической структурой сохранялась и преобладала значительно дольше, чем в западной Евразии. Пока менее чем 4 тыс. лет назад по степи не прокатились волны миграций, которые привнесли в регион одомашненные породы лошадей с последующим появлением нового снаряжения для верховой езды и колесницы со спицами. Также эти миграции способствовали распространению проса в качестве сельскохозяйственной культуры.
Источник: Allentoft, M.E., Sikora, M., Refoyo-Martínez, A. et al. Population genomics of post-glacial western Eurasia. Nature 625, 301–311 (2024). https://doi.org/10.1038/s41586-023-06865-0
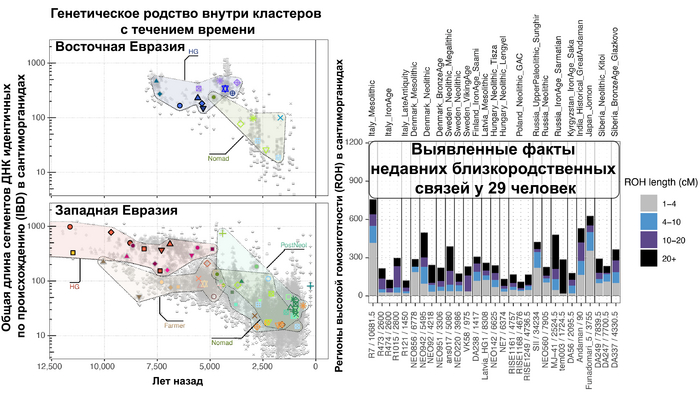
Закономерности распределения сегментов ДНК идентичных по происхождению между людьми, продемонстрировали снижение родства внутри кластеров с течением времени как в западной, так и в восточной частях Евразии. Что согласуется с увеличением эффективного размера популяции в течение этого времени. Однако наблюдаются заметные демографические различия между западной и восточной Евразией. Если на западе во время перехода к неолиту около 9000-6000 лет назад, люди с родословной анатолийских земледельцев демонстрируют снижение доли сегментов ДНК идентичных по происхождению, по сравнению с охотниками-собирателями, то на востоке генетическое родство оставалось высоким примерно до 4000 лет назад, что согласуется с гораздо более длительным существованием небольших локализованных групп охотников-собирателей.
Помимо этого, анализ сегментов гомозиготности (ROH) не выявил значительного числа близкородственных связей, только 29 из 1396. При этом каких-то пространственно-временных или культурных закономерностей в распространенности близкородственных связей обнаружено не было. Зато авторы задокументировали первый случай однородительской дисомии по древнему геному (когда человек получает две копии хромосомы или части хромосомы от одного родителя и ни одной копии от другого родителя). Эта особенность обнаружена у представителя из Темясовских курганов позднесарматской культуры возрастом около 1700 лет.
Также были выявлены люди с различной степенью родства.
Итоги
Появление и сокращение генетического разрыва
После максимума последнего оледенения около 25-19 тыс. лет назад во время расселения древних людей по Евразии, установилось чёткое генетическое разделение между востоком и западом по линии от Черного моря до Балтийского, что отражено и в археологических данных. Это разделение сохранялось на протяжении всего мезолита и неолита. В Восточной Европе распространение земледелия происходило с опозданием примерно на 3000 лет, и эта задержка может быть связана с особенностями окружающей среды, поскольку регионы к востоку от условной границы имеют более континентальный климат и более суровые зимы, возможно, менее подходящие для ближневосточных методов ведения земледелия. На этих территориях сохранились высокоразвитые сообщества охотников-собирателей со стабильными, сложными, а иногда и укрепленными поселениями, с обменом на большие расстояния и крупными кладбищами. Рацион, включающий пресноводную рыбу, зафиксирован как по изотопным данным из этого исследования, так и по анализам остатков липидов в керамике.
В северных лесных районах этой пограничной зоны сообщества охотников-собирателей сохранялись вплоть до появления культуры шнуровой керамики около 5 тыс. лет назад, тогда как в южных и восточных степных регионах охота и собирательство со временем были дополнены животноводством с крупным рогатым скотом и овцами и, возможно, коневодством в Центральной Азии, но этот факт ещё под вопросом. Некоторые из этих групп, такие как представители хвалынской культуры Северного Кавказа и Среднего Поволжья, стали свидетелями появления мужских сообществ, вовлеченных в широкомасштабные обменные связи на основе медных изделий с Восточной и Центральной Европой, а также Кавказом. Поселения в то время были приурочены в основном к равнинным поймам и долинам рек, тогда как степной пояс оставался в значительной степени неиспользованным.
Окончательное исчезновение этой генетической, экономической и социальной границы было вызвано событиями, развернувшимися в степном регионе. Здесь археологически можно наблюдать две временные фазы технологических инноваций: широкое распространение колесных повозок, запряженных волами, около 5500 лет назад и более позднее развитие колесниц со спицами и верховой езды.
Начиная ещё с первой фазы инноваций, в сочетании с возможными изменениями условий окружающей среды, степь стала выгодной экономической зоной, которая позволила группам по типу ямной культуры эксплуатировать степь в качестве кочевых степных скотоводов около 5000 лет назад. А поселения эпохи энеолита вдоль речных долин были заменены этой новой формой мобильной экономики на основе скотоводства, которая окончательно разрушила генетическую границу, существовавшую в предыдущие тысячелетия.
Далее, после второй фазы инноваций около 4 тыс. лет назад, изобретение боевых колесниц и использование проса в качестве продовольственной культуры позволили андроновцам и родственным им группам окончательно распространиться на восток в Среднюю Азию и за ее пределы, оставив глобальное наследие в виде распространения индоиранских языков.
Это исследование внесло свою лепту в наши знания о прошлом, особенно касательно степных миграций.
Во-первых, авторы выявили до сих пор неизвестный источник родословной, который способствовал появлению степных скотоводов, на его роль хорошо подходили охотники-собиратели Среднего Дона, как люди из могильника мариупольского типа Голубая Криница раннего энеолита на юге Воронежской области, возрастом около 7300 лет.
А во-вторых, было определено, что степная родословная, которая распространилась по Европе, благодаря представителям культуры шнуровой керамики, сформировалась в результате культурного и генетического слияния степных групп, родственных ямной культуре и культуре шаровидных амфор. При этом благодаря своему смешанному культурному и генетическому происхождению, носители культуры шнуровой керамики практиковали смешанную экономику, используя различные стратегии выживания в различных условиях. Такая гибкость в значительной степени способствовала их успеху в расселении и адаптации к очень разным экологическим и климатическим условиям за очень короткий период времени.
Источник: Allentoft, M.E., Sikora, M., Refoyo-Martínez, A. et al. Population genomics of post-glacial western Eurasia. Nature 625, 301–311 (2024). https://doi.org/10.1038/s41586-023-06865-0
Два параллельных мира Европы и война, которой почти 5 тысяч лет
То что будет рассказано в этой статье для части моих читателей, которые мало знакомы с древней историей человечества, будет неожиданностью и возможно даже приведёт к внутреннему сопротивлению и не согласию. Так уж устроен наш мозг. Одним из основных факторов, препятствующих усвоению любой новой информации, является наше стремление сохранять привычный порядок вещей. Мозг привык к выработанным в течении всей нашей жизни шаблонам, схемам и стереотипам, которые помогают ему более быстро обрабатывать информацию. Если новые данные противоречат этим устоявшимся схемам, мозг будет сопротивляться их усвоению в попытке сохранить старый порядок. Разрыв шаблонов и слом привычной картины мира всегда крайне болезненно воспринимается всеми людьми, так как часто мешает им приспосабливаться к существующей действительности. По этой же самой причине явные различия в восприятии окружающего нас мира между людьми нередко приводили их не только к конфликтам, но и беспощадным войнам.
Война Атлантиды с Гипербореей является одной из наиболее "запретных тем" современной истории. Ведь, если признать ее существование, то рано или поздно откроются и причины, по которым война возникла и привела к гибели этих развитых цивилизаций. Согласно общепринятой точке зрения, сложившейся на основе древних легенд место расположения как Гипербореи, так и Атлантиды точно не известны. Гиперборею в основном связывают с севером Евразии или Уралом, а Атлантиду - с более южными районами (Атлантикой и Средиземноморьем), размещая ее где-то от Крита, Гибралтара и до Британских островов. По древним преданиям обе великие цивилизации обладали магическими знаниями. Пантеон гиперборейских богов и магических знаний был связан с белым цветом (снегом и севером), а пантеон атлантов с черным цветом (черноземом, черной землей и югом). Именно с тех давних пор пошли названия белая и черная магия. Согласно древним мифам Гипербрейцы и Атланты существовали вместе с древних времен, но атланты занимали более плодородные южные земли, в которых всегда была выше плотность населения, конкуренция и борьба за свое выживание. По этой причине менталитет обеих цивилизаций очень сильно различался. Для гиперборейцев в борьбе за выживание более важными аспектами жизни были коллективизм и философия (сейчас бы это назвали академической наукой), а для атлантов в борьбе за выживание на первом месте всегда был индивидуализм, прагматизм и поэтому большая агрессивность. Именно конкурентность за жизненные ресурсы, жесткая индивидуальная борьба между отдельными особями в популции за их обладание всегда порождают агрессивность. Наоборот, переход к более коллективным формам деятельности всегда снижают конкурентность за ресурсы и агрессивность, и увеличивают взаимопомощь и сотрудничество, а не вражду. Это характерно не только для человека, но и для всего мира живых существ. По некоторым древним легендам, после того как атланты сравнялись в своих знаниях и могуществе с цивилизацией гиперборейцев они развязали войну.
Два параллельных мира Европы и война, которой почти 5 тысяч лет
Но это все легенды, мифы и предания, а что же говорит наука?
Начну с основных научных фактов, которые были получены относительно недавно и подтверждены не только археологами, но и генетиками после расшифровки ДНК древних людей, живших более 5 тысяч лет назад на Русской равнине и в современной Европе.
Факт №1. Современные жители Европы, как западной, так и восточной, на 80% происходят всего от двух человеческих племен, которые еще 5-10 тысяч лет назад проживали только на Русской равнине. Позже ученые придумали этим двум древним племенам название - индоевропейцы. Остальную же Европу в те далекие времена заселяли совершенно другие народы, которые были неродственными по Y-ДНК индоевропейцам.
Оказалось, что Русская равнина была заселена довольно неравномерно, а два племени, которые жили на ней имели родственные геномы R1 (R1a и R1b), но занимали совсем разные ниши. Южные степные районы России, от Южного Урала до Дона населяли племена с Y-ДНК гаплогруппой R1b (их с легой руки А. Клесова стали называть эрбины). А вот более северные лесостепные и лесные районы на Русской равнине населяли племена с другой Y-ДНК гаплогруппой - R1a. Довольно долго оба племени были соседями и почти не конфликтовали. Носители Y-ДНК R1b известны в первую очередь по Ямной культуре или предшествовавших ей культурам: Среднестоговой, Самарской и Хвалынской, а эраны или арии (носители Y-гаплогруппы R1a) по культуре Шнуровой керамики или Боевых топоров. Не наводит ли вас на ассоциации с северянами и южанами, атлантами и гиперборейцами? Если нет, то тогда идем дальше.
Сейчас народы-носители R1b населют в основном Западную Европу. Более всего Y-гаплогруппа R1b распространена вдоль Атлантического побережья и особенно среди англо-саксов, достигая в этих популяциях максимума (до 90%) на Британских островах. Народы-носители R1a преимущественно живут в Восточной Европе, достигая максимума в Польше, Украине, России и Беларуси (в отдельных районах до 60-70% от общей численности). И те и другие этно-генетические популяции народов возникли изначально где-то в центральной Сибири, но на рубеже 9-10 тысяч лет назад стали проникать в Европу, но разными маршрутами. Носители R1a пришли туда первыми и сейчас считаются условными автохтонами (коренными жителями) Русской равнины. Они двигались северным путем, а носители R1b южным, через Среднюю Азию и Иран. Ну теперь мне кажется ассоциации с атлантами и гиперборейцами, южанами и северянами начинают возникать у читателей? Если нет, то тогда идем дальше.
Факт №2. Примерно 5 тысяч лет назад во времена бронзового века степь стала высыхать и стада копытных животных начали все больше и больше мигрировать на север. Ресурсов для жизни у южных племен (R1b) становилось все меньше и они попытались атаковать северян (R1a) и захватить их земли, однако, получили достойный отпор. Началась длительная кровопролитная война за ресурсы, которая продолжалась несколько столетий. В результате южане были разгромлены и покинули места своего проживания, двинувшись на запад. Знаментиая американская ученая-археолог Мария Гимбутус создавшая так называемую "Курганную теорию" впервые рассказала общественности о жестокости эрбин, которые в процессе захвата Западной Европы (2900-2000 гг. до нашей эры) практически полностью уничтожили все мужское население Старой Европы. Лишь очень небольшой его части удалось тогда выжить в наиболее труднодоступных местах - горных районах Балкан и в Скандинавии или же вступить в союз с ариями (носителями R1a) дав начало предкам славян и других восточноевропейских народов.
В течение последующих тысячелетий борьба между этими двумя этно-генетическими группами народов Европы происходила постоянно и очень жестко, а граница все время сдвигалась то на восток, то на запад. Однако, как показала история, агрессорами всегда были только одни - эрбины или западные европейцы (начиная с римлян, потом крестоносцев, вплоть до Наполеона и Гитлера). Вероятно, естественный отбор закреплял мутации агрессивного поведения у эрбин и в дальнейшем от них пострадали не только европейцы, но и многие народы Азии, Америки и Африки.
Два параллельных мира Европы и война, которой почти 5 тысяч лет
Танковое противостояние у КПП «Чарли», известное также как Танковое противостояние на Фридрихштрассе или Инцидент у КПП «Чарли» — эпизод Берлинского кризиса 1961 года
Интересно, что во времена СССР граница между двумя этно-генетическими группами народов Европы (R1b и R1a) совпала с границами двух военных блоков (НАТО и Варшавский договор), как бы это ни казалось парадоксально (в то время о различиях в ДНК западных и восточных европейцев ученые еще не догадывались). Сама же разделительная линия между этими враждовавшими блоками тогда проходила по нейтральным странам - начиная от бывшей Югославии на юге, через Австрию и Швейцарию в центре Европы и до Скандинавии и Финляндии на севере. Интересно, что во всех нейтральных странах Европы население было либо смешанным по Y-ДНК гаплогруппам, либо имело совсем другие ДНК-маркеры, не свойственные ни западноевропейцам, ни восточноевропейцам. Даже сама Германия была разделена на ГДР и ФРГ казалось бы не случайно, а буд-то специально по Y-ДНК маркерам (хотя тогда об этом никто не знал). Дело в том, что в ГДР и во всей Восточной Германии как у русских преобладают восточноевропейские геномы (R1a), а в Западной Германии (тогдашней ФРГ) - западноевропейские генотипы (R1b). Сейчас эрбины (носители гаплогруппы R1b) вновь агрессивно сдвигают свою границу все дальше и дальше на восток, объясняя все это национальной безопасностью и борьбой за права других народов. Они включили в число своих сателлитов многие восточноевропейские народы, идентичные по генетике русским. Уж не используют ли они опять свою "черную магию" или что-то еще? Более того, западные политики кулуарно продвигают полностью антинаучные теории об отличии других восточных европейцев от русских и даже особой агрессивности последних. Как говорится в известной поговорке: "С больной головы да на здоровою", то есть когда люди не в состоянии принять свои ошибочные устремления все время ищут виноватых среди других.
Если Вам понравилась статья подписывайтесь на канал, делитесь ею в социальных сетях и ставьте лайк (👍 - палец вверх)
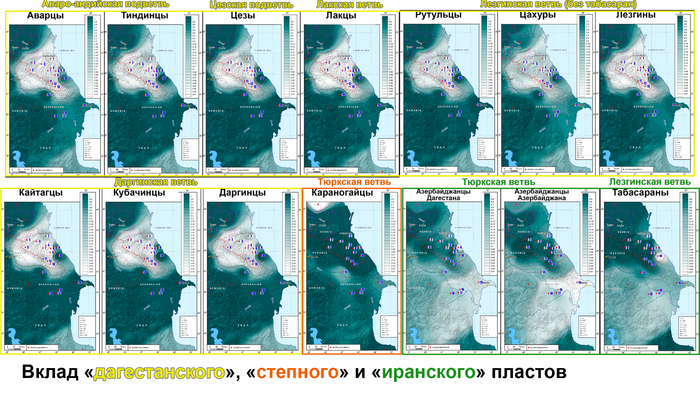
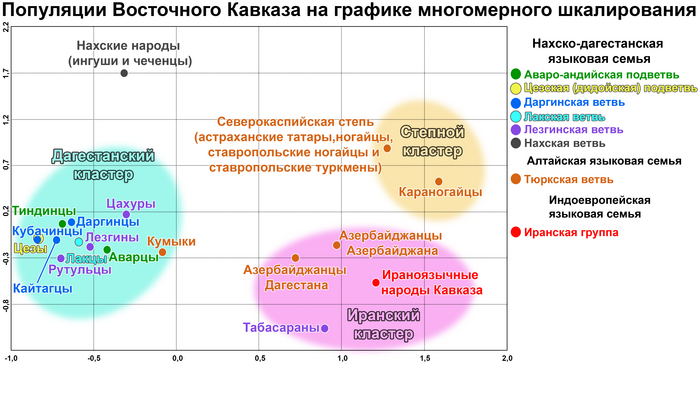
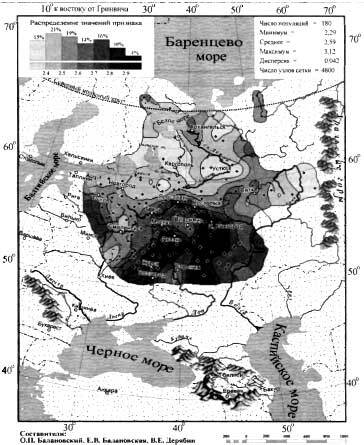
Положение 22 народов на графике анализа главных компонент
На графике анализа главных компонент четко выявляется 5 кластеров, которые показывают, что сходство аутосомных геномов народов Восточного Кавказа очень слабо сочетается с их языками.
1/2
Почти все кластеры включают народы, говорящие на языках не только разных ветвей, но даже разных лингвистических семей. Первая главная компонента четко делит исследованные народы на степные и все остальные, а вторая демонстрирует длинную цепь всех остальных геномов — от кубачинцев до курдов.
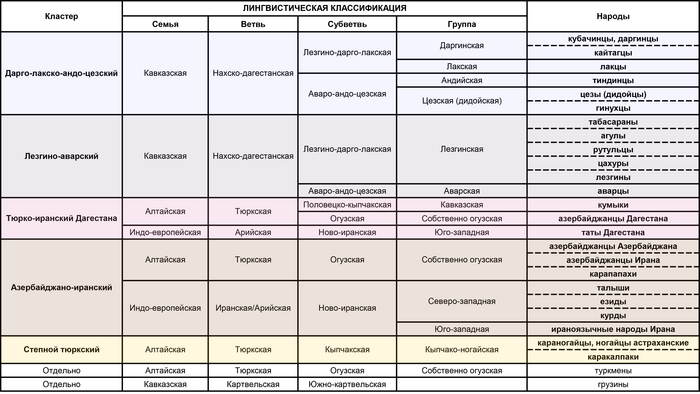
Дарго-лакско-андо-цезский кластер
В этот кластер вошли: кубачинцы, даргинцы, кайтагцы, лакцы, тиндинцы, цезы и гинухцы. Однако стоит отметить, что кубачинцы заметно отклонены от основного скопления, сильнее чем цезы и гинухцы.
Лезгино-аварский кластер
Объединяет представителей шести этносов из лезгинской и аварской ветвей нахско-дагестанских языков. При этом аварцы крайне неоднородны и разбросаны на графике вдоль всего лезгино-аварского кластера и уходят за его пределы к представителям дарго-лакско-андо-цезского кластера. А лезгины, наоборот, тянутся к татам и азербайджанцам Дагестана, тюрко-иранского кластера Дагестана.
Тюрко-иранский кластер Дагестана
Этот кластер объединил геномы ираноязычных татов с тюркоязычными кумыками и азербайджанцами Дагестана. При этом часть кумыков попадает в лезгино-аварский кластер, хотя в целом геномы всех трех этносов тяготеют к азербайджано-иранскому кластеру. Стоит отметить, что в Дагестане татов было принято записывать как «азербайджанцев», поэтому сходство геномов татов и азербайджанцев Дагестана не удивляет.
Азербайджано-иранский кластер
Кластер также объединил геномы тюркоязычных и ираноязычных народов, а именно карапапахов, азербайджанцев Азербайджана и Ирана с талышами, курдами, езидами и другими группами Ирана из предыдущих исследований). При этом карапапахи демонстрируют небольшой сдвиг в сторону народов Евразийской степи. А геномы туркмен демонстрируют своеобразный мост между азербайджано-иранским и степным кластерами. С другой стороны, положение грузин на графике связывает азербайджано-иранский и тюрко-иранский кластеры Дагестана.
Степной кластер
Объединил караногайцев Дагестана с другими степными народами Прикаспийской степи и Закаспия, а именно астраханских ногайцев с каракалпаками.
Карта на графике позволяет увидеть взаимодействие трех генетических пластов в ареале Восточного Кавказа. С юга распространяется влияние Ирана, захватывающее весь Азербайджан и проникающее в Дагестан вдоль берега Каспийского моря. А Дагестан в свою очередь сохраняет генетическое своеобразие автохтонного населения восточной окраины Северного Кавказа, тогда как с севера отмечено более позднее влияние евразийской степи. При этом эти три генетических пласта слабо связаны с тремя языковыми семьями на которых говорят исследованные народы.
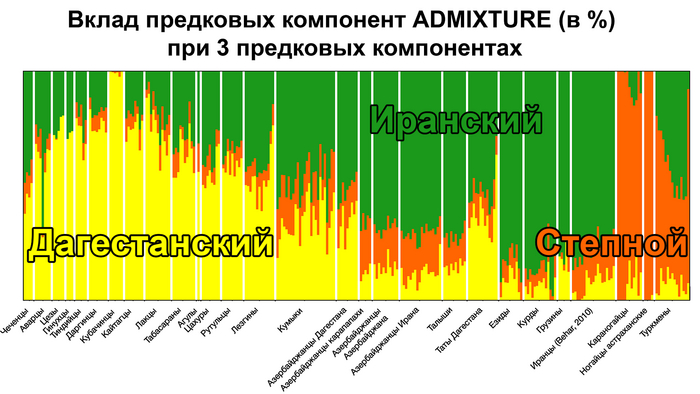
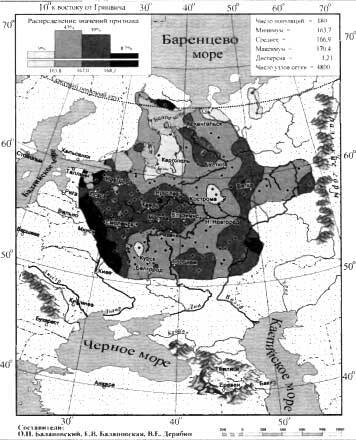
Вклад предковых компонент (ADMIXTURE) в генофонды народов Восточного Кавказа
В работе из всех возможных моделей от 2 до 20 предковых компонент наиболее информативными, по мнению учёных, оказались модели с 3, 10 и 20 предковыми компонентами.
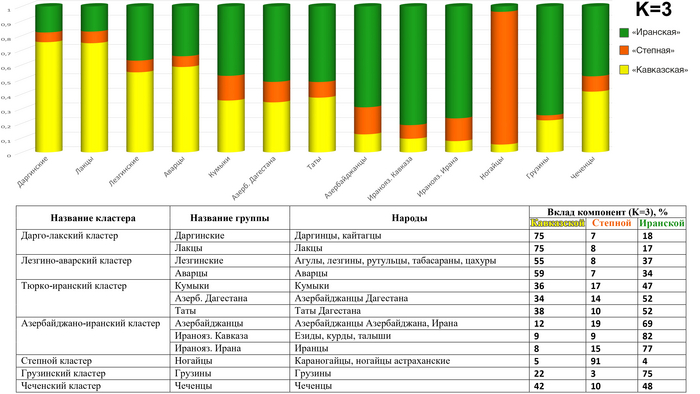
Модель 3 предковых компонент
Результаты моделирования могут служить подтверждением гипотезы «трех пластов» генофонда Восточного Кавказа: оранжевый цвет, характерный для караногайцев, отражает гипотетический «степной» генетический пласт; зеленый цвет, ярко выраженный у курдов и ираноязычных иранцев, — «иранский» пласт; желтый цвет — «кавказский» пласт, преобладает в геномах народов Дагестана и, демонстрирует вклад кавказоязычного населения региона. В целом геномные профили предковых компонент соответствуют и кластерам в анализе главных компонент.
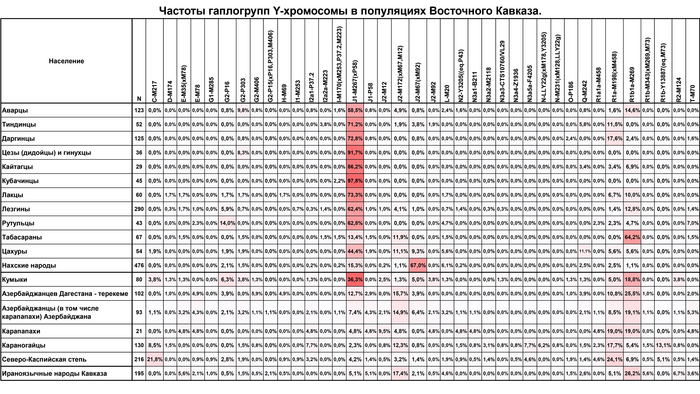
«Кавказский» вклад в геномах дарго-лакского кластера составляет 75%; в лезгино-аварском — чуть более половины генофонда; в тюрко-иранском — около трети, в азербайджанско-иранском — около 10%. По мере убывания «кавказского» пласта столь же резко возрастает «иранский»: 17%, 35%, 50%, 75%. «Степной» пласт оказывается мощным только в степном кластере (91%): у кавказоязычных народов он составляет всего 7–8%, а у тюркоязычных — 14–19%.
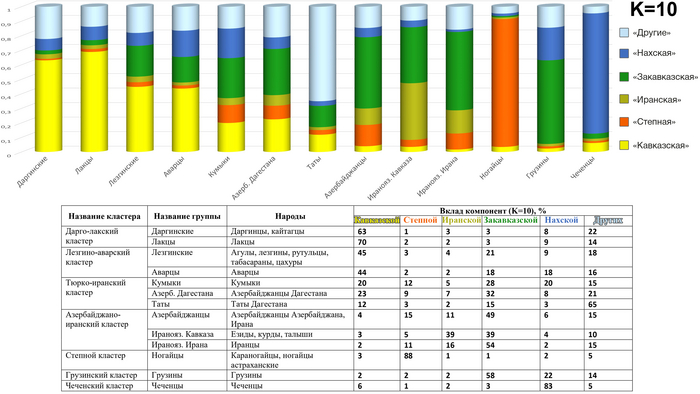
Модель 10 предковых компонент
При моделировании 10 предковых компонент для той же совокупности геномов наблюдается сохранение трех исходных предковых компонент («степной», «иранской» и «кавказской»). Однако их соотношение частично меняется за счет появления новых компонент — «нахской», которая преобладает у чеченцев, и «закавказской», составляющей у грузин более половины их аутосомного генофонда. «Нахская» предковая компонента забирает на себя в основном вклад от «кавказского» пласта, а «закавказская» от «иранского». При этом учитывая, что вклад «закавказской» предковой компоненты в геномы ираноязычного населения Ирана около 54%, что практически равно её вкладу в геномы самих грузин (58%), можно предположить, что она отражает древний западноазиатский пласт в генофондах Кавказа и Закавказья.
А новые компоненты, которые вносят небольшой вклад в геномы других народов, объединены в одну предковую компоненту - «Другие». К ним отнесены предковые компоненты малочисленных этносов (кубачинцев, цезов, гинухцев и тиндинцев), обычно отражающие близкородственные геномы в их пределах, а также особые варианты предковых компонент у части геномов татов и туркмен.
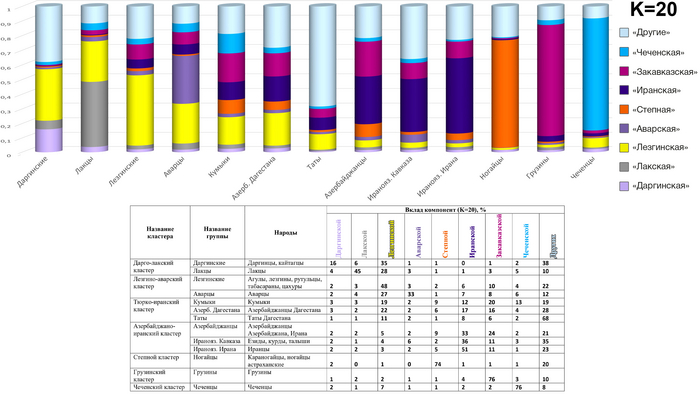
Модель 20 предковых компонент
При моделировании 20 предковых компонент, также появляются новые компоненты, которые отражают генетическое своеобразие народов даргинской и лезгинской групп, аварцев и лакцев. Из них наибольший вклад в геномы других народов Дагестана вносит «лезгинская» компонента, составляя треть генофондов дарго-лакского и аварского кластеров, половину лезгинского и пятую часть генофонда кумыков.
Однако появление в этой модели «лезгинской», «лакской» и «даргинской» предковых компонент, которые вписываются в общий «кавказский» генетический пласт, отраженный желтым тоном в предыдущих моделях под названием «кавказская» предковая компонента, даёт повод для его переименования в «дагестанский» генетический пласт, что более точно отражает его суть.
Три генетических пласта
Анализы различными дополняющими друг друга методами как гаплогрупп Y-хромосомы, так и аутосомного генофонда Восточного Кавказа выявляют наличие трех генетических пластов, из которых «иранский» связан с многочисленными волнами иранского населения, «степной» — с поздним влиянием популяций евразийских степей, а «дагестанский» — с древним населением, сформировавшимся в ареале современного Дагестана. Мощность каждого из этих генетических пластов варьируется в разных группах населения Восточного Кавказа, однако хорошо сочетается с географией.
«Степной» генетический пласт составляет основу генофондов многих народов Каспийского региона (астраханских ногайцев, каракалпаков, туркмен) и, хотя он ограничен только небольшим ареалом евразийской степи на севере региона, он затрагивает многие народы на значительном расстоянии. Однако максимума генофонда он достигает лишь у астраханских ногайцев и караногайцев. Тогда как в остальных геномах региона его вклад варьируется в пределах от 7% у народов Дагестана до 19% в популяциях Азербайджана.
«Иранский» генетический пласт у популяций Восточного Кавказа играет очень важную роль, распространяясь в регион с Иранского нагорья он прослеживается у всех исследованных народов Азербайджана, составляя три четверти генофонда и почти полностью растворяется среди народов Дагестана от лезгинских групп на юге и кумыков с татами в Прикаспии, к центру и северу, составляя в среднем около трети генофонда у народов Дагестана, если не считать ногайцев.
«Дагестанский» генетический пласт представляет собой древний генетический пласт, географически расположенный между другими пластами и составляет основу всех народов Дагестана, и в целом больше половины генофонда кавказоязычных народов от 52 до 100% (в среднем 66%), несмотря на то, что при увеличении числа предковых компонент практически у всех изученных народов Дагестана появляется собственная предковая компонента.
Стоит отметить, что Прикаспийский регион является перекрёстком и местом встречи всех трех пластов: у кумыков, азербайджанцев Дагестана и татов половину генофонда составляет «иранский» пласт, треть — «дагестанский» и около 15% «степной».
Генетика и лингвистика
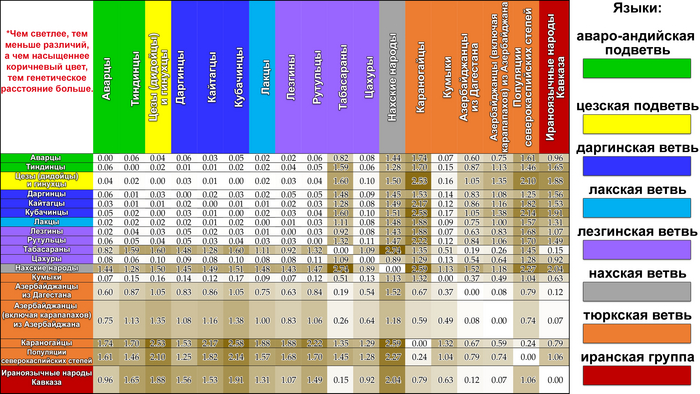
В отличие от географии, связь между генетикой и языками очень слаба. В основном это обусловлено переходом на тюркские языки народов Восточного Кавказа (кумыков и азербайджанцев), во многом сохранивших при этом «дотюркский» генофонд.
Также заметно явное влияние окружающих народов и на геномы народов, сохранивших свой язык. Так, «дагестанский» генетический пласт у татов Дагестана достигает 38%, тогда как у других ираноязычных народов Восточного Кавказа (курдов, езидов и талышей) — всего 9%. Кстати у грузин, представителей картвельского языка, во всех моделях, менее 19 предковых компонент, генофонд в среднем на три четверти формируется «иранским» пластом.
Выводы
Три пласта генофонда Восточного Кавказа неодинаковы и, скорее всего, возникли в разные периоды истории популяций. Также взаимодействие этих трех генетических пластов обусловлено географическими особенностями региона и лишь косвенно связано с лингвистикой.
«Степной» пласт встречается только на севере региона среди караногайцев и отражает самую последнюю миграционную волну тюркоязычных народов из евразийских степей в средние века. А «дагестанский» с «иранским» образуют два противоположных полюса и вносят наибольший вклад в генофонд Восточного Кавказа.
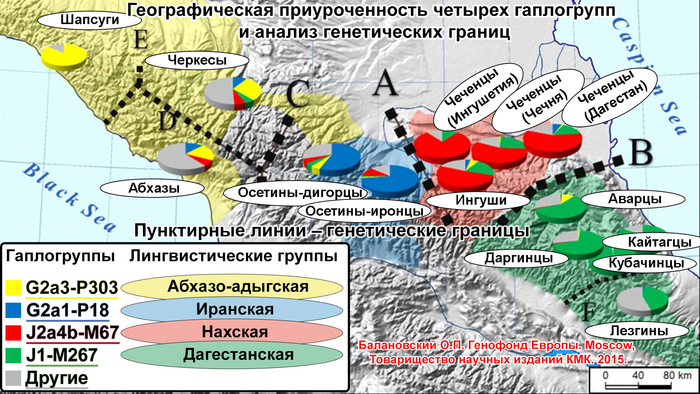
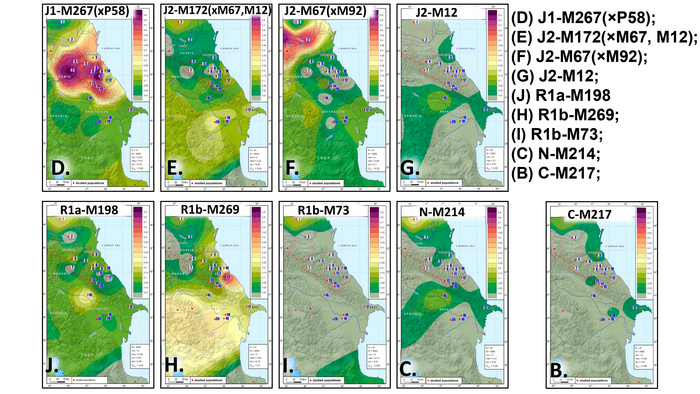
«Иранский» пласт сформирован многочисленными волнами иранского населения в ареале Восточного Кавказа. Этот пласт показывает генетическое сходство азербайджанцев, табасаран и ираноязычного населения Кавказа с населением Западной Азии, что может быть связано с серией древних миграционных волн. Эта закономерность является результатом вклада гаплогрупп J2-M172 (×M67, M12) и R1b-M269, которые, возможно, были занесены в регион миграциями западноазиатских племен на протяжении всей его истории, в основном говорившим на иранском языке.
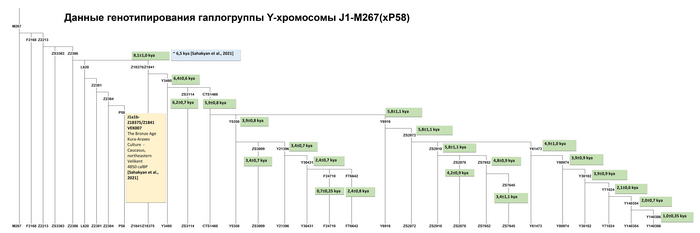
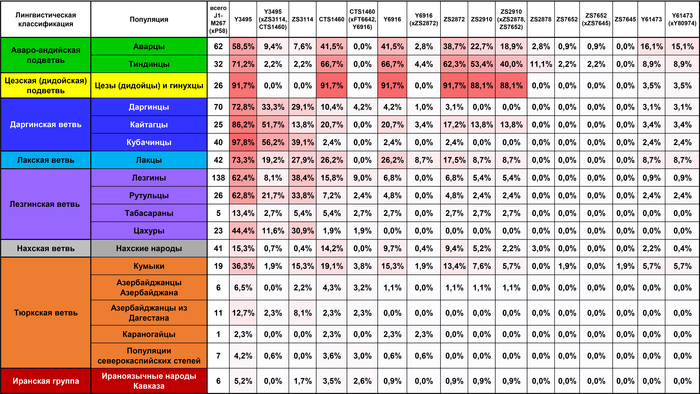
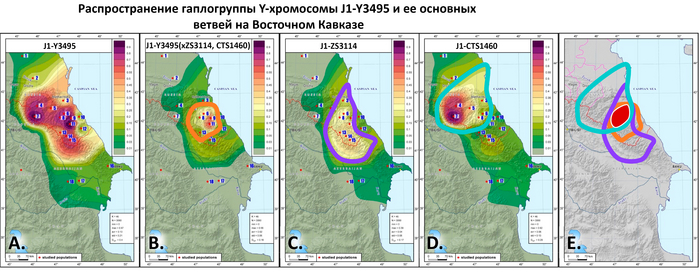
«Дагестанский» пласт отражает генофонд древнего населения Северного Кавказа. При моделировании большого числа предковых компонент у большинства народов Дагестана выявляется собственная предковая компонента, но все этносы Дагестана объединяет мощный общий древний генетический пласт. По данным анализа гаплогрупп Y-хромосомы он в значительной степени сформирован линией J1-Y3495 (6,5 ± 0,6 тыс. лет назад), появлению которой, возможно, предшествовало сокращение численности населения, за которым последовал её рост. Анализ филогеографии J1-Y3495 позволил проследить происхождение этой линии в центральной части горного Дагестана, от предкового населения, говорившего на пра-нахско-дагестанском языке. А раскол в генофонде около ~6 тыс. лет назад может быть связан с ростом популяции, рассредоточением сообществ и их длительной изоляцией в горах. Это подтверждается расщеплением этой гаплогруппы в популяциях аваро-андо-цезской ветви ~4-5 тыс. лет назад.
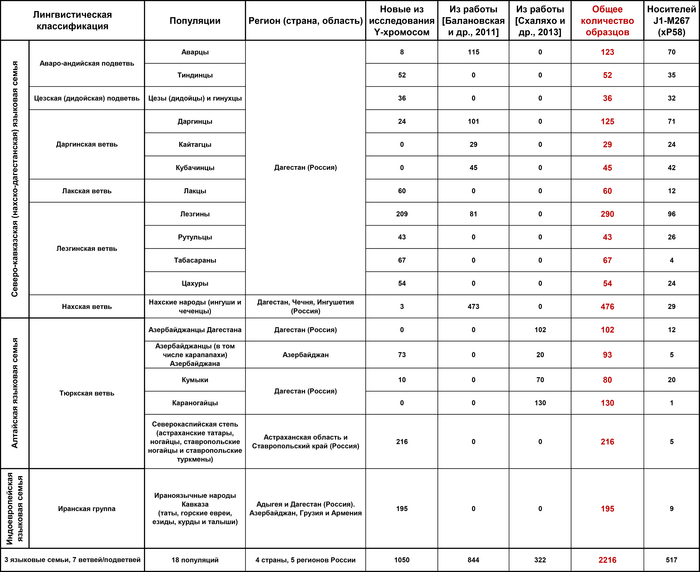
Помимо этого, одной из целей работы по анализу аутосомного генофонда был поиск таких групп этносов, которые могут надежно представлять Восточный Кавказ в евразийском масштабе. Два независимых метода указывают на необходимость включения четырех таких групп:
1) даргинцы и лакцы;
2) аварцы, лезгины, табасараны, агулы, рутульцы и цахуры;
3) кумыки, таты и азербайджанцы Дагестана;
4) азербайджанцы и талыши Азербайджана.
Их комплекс создает корректное представительство многообразия Восточного Кавказа.
Поскольку «степной» вклад геномов караногайцев в данном случае невелик, их геномы правильнее рассматривать в контексте генофонда евразийской степи или же ее каспийского сегмента.
Источники:
Балановская, Е. В., Горин, И. О., Петрушенко, В. С., Пономарёв, Г. Ю., Белов, Р. О., Почешхова, Э. А. и др. Роль кавказского, иранского и степного населения в формировании многообразия аутосомного генофонда Восточного Кавказа // Вестник РГМУ. 2023. №3. С. 36–45. DOI: https://doi.org/10.24075/vrgmu.2023.017
Примечание: текста очень много, тема сложная и состоит из двух постов с полным названием "Генофонд народов Восточного Кавказа на основе гаплогрупп Y-хромосомы и аутосомных данных". Выводы к обеим частям во второй части, после аутосомных данных.
Примечание к видео: там, где я говорю: "включая линии" P58, M67, M12, M458 и другие субклады, ошибка! Правильно "без этих линий" Пример: J2(×M67, M12) - надпись в скобках обозначает отсутствие этих мутаций, т.е., не включая этих ветвей. (Коррекцию предложил внести один из соавторов работы – Насиб Искандаров)
Вступление
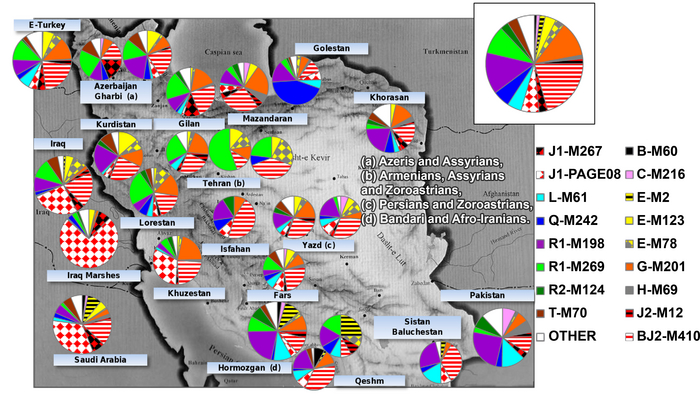
Восточный Кавказ, который охватывает современные Дагестан и Азербайджан, является важным сухопутным мостом, соединяющим Европу с Передней или Западной Азией (далее буду употреблять термин — Западная Азия). Одним из путей через Кавказ является Большой Кавказский проход, прибрежная полоса которая проходит между горами Большого Кавказа и Каспийским морем от Дербента до Сумгаита. Эта территория была населена представителями рода людей, носителями олдувайской культуры, ещё около 2 миллионов лет назад.
*Не предки современных людей
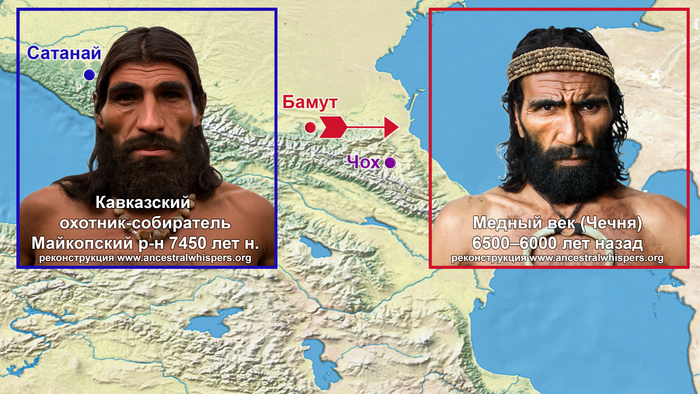
А что касается людей современной анатомии, то древнее поселение Чох в нынешнем Дагестане является ключевым памятником культур мезолита, неолита и бронзового века, которые раньше существовали в горных районах Северо-Восточного Кавказа. Данные недавнего радиоуглеродного датирования свидетельствуют о том, что одомашнивание животных, выращивание сельскохозяйственных культур и изготовление керамики в этом регионе началось не позднее конца 7-го или начала 6-го тысячелетий до нашей эры.
Экономическое развитие в период неолита (земледелие и животноводство, появление керамики, ткачества, новых технологий камнеобработки) привело к быстрому росту населения и колонизации новых территорий. На территории современного Азербайджана развитые неолитические общества возникли на заре 6-го тысячелетия до нашей эры как преемники неолитических обществ Юго-Западной Азии. Открытие металлов в эпоху энеолита (~7 тыс. лет назад) стимулировало развитие технологий. Бронзовый век (6-4 тыс. лет назад) ознаменовался появлением новых методов обработки металла, экономическими и социальными изменениями, включающими социальное неравенство и племенные объединения, а также торговыми отношениями с населением Юго-Восточной Европы и Западной Азии. А уже в среднем бронзовом веке на территорию Восточного Кавказа прибыли кочевники из евразийских степей, привнеся в регион традиции курганных захоронений.
В VII–VI веках до нашей эры на юг региона через Дербентский перевал проникли ираноязычные скифы, которые образовали военно-политический союз, известный как Ишкуза, или Скифское царство, на землях современного Западного Азербайджана и Северо-Западного Ирана. Позже эта территория была присоединена к Мидийскому царству. В конце первого тысячелетия до нашей эры из союза племён в северной части региона возникло государство Кавказская Албания. У него была своя собственная письменность, и его население приняло христианство в IV веке нашей эры. Кавказская Албания в значительной степени состояла из автохтонных кавказских племен, говоривших на лезгинских языках нахско-дагестанской семьи, хотя ее господство распространялось и на некоторые иранские племена.
Территория современного Дагестана была колыбелью таких ранних государств, как Лакз, Табасаран, Кайтагское уцмийство, Сарир, Царство Гумик и др., возникших в первой половине 1-го тысячелетия нашей эры.
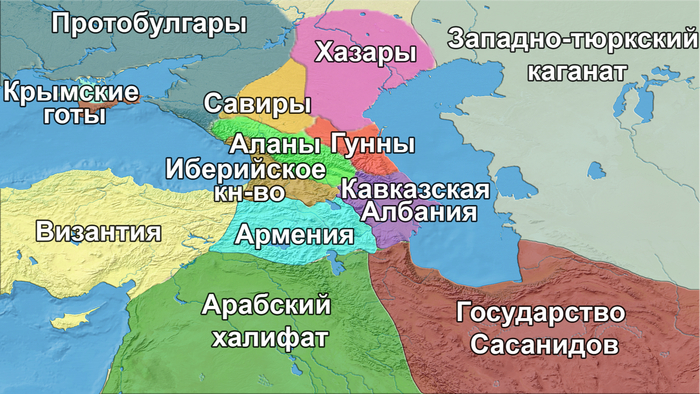
В течение первых нескольких столетий нашей эры на Кавказ прибывали другие иранские племена, включая сарматов, маскутов и алан. А Южный Кавказ подвергался набегам гуннов, савиров и хазар, приходившим с севера через Дербентский перевал в раннем средневековье (между концом IV и VIII веками).
В VII веке нашей эры север равнинного Дагестана был захвачен развивающимся Хазарским каганатом, в то время как юг региона был в составе государства Сасанидов с IV века.
В VII–IX веках Восточный Кавказ контролировался Арабским халифатом, который проводил агрессивную миграционную политику, приведшую к исламизации местного населения.
Вторжения тюркоязычных сельджуков, основателей могущественной империи, охватывающей Центральную и Западную Азию, а также Южный Кавказ, начались в XI веке, когда захватчики продолжали экспансию из Средней Азии на Восточный Кавказ, включая Дербент.
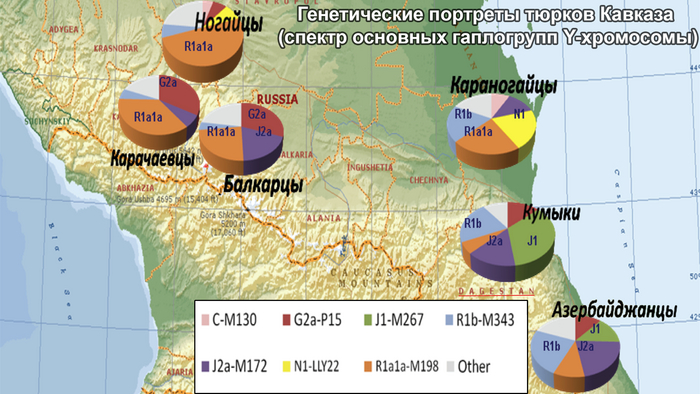
Тюркские народы из Половецкой или Кыпчакской степи, мигрировали на Южный Кавказ в XI- XII веках, а их потомки, возможно, внесли свой вклад в возникновение карапапахской группы среди азербайджанцев.
Другими тюркоязычными народами Восточного Кавказа являются кумыки и ногайцы. Единого мнения о происхождении кумыков нет. Преобладающая гипотеза прослеживает их происхождение от местного автохтонного населения, имевшего тесные этнокультурные контакты с чужеземными тюркскими племенами, такими как савиры, хазары или кыпчаки (половцы). В свою очередь, ногайцы были кочевниками поздней миграционной волны; большинство из них осело в Дагестане и представлено караногайцами.
После Монгольских завоеваний, с XIV века, Дагестан снова попал под исламское влияние.
Местные средневековые княжества того времени просуществовали до XIX века, когда они были поглощены Российской империей.
Южные регионы Восточного Кавказа находились под иранским влиянием в течение XIV-XIX веков; они были включены в состав Российской империи после русско–персидских войн, когда в 1813 и 1828 годах были подписаны мирные договоры. Позже эти территории вошли в состав Азербайджана. Это очень сокращённая история демонстрирует, что на генофонды автохтонных популяций Восточного Кавказа оказали влияние как массовые древние миграционные волны ираноязычных народов, так и более поздние миграции евразийских тюркоязычных народов. Богатая история региона отражена также и в языковом разнообразии Восточного Кавказа.
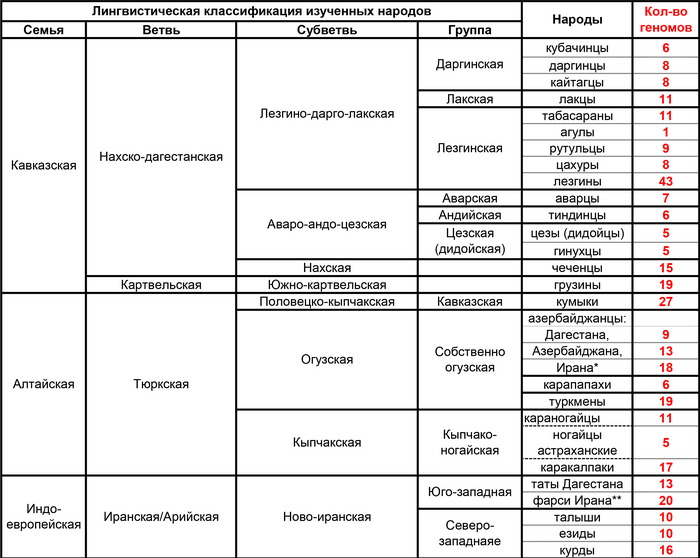
Языки региона включают нахско-дагестанские языки (на которых говорят более 30 этнических и этнографических групп), тюркские языки алтайской языковой семьи (на которых говорят азербайджанцы, кумыки и караногайцы) и иранские языки индоевропейской языковой семьи (на которых говорят таты, горские евреи, езиды, курды и талыши). Такое значительное разнообразие популяций Восточного Кавказа и их необычайная структура является серьезным препятствием в изучении генофонда народов, проживающих в регионе.
Некоторые предыдущие популяционные исследования указывают на сходство между генофондами азербайджанцев и их соседей на Кавказе и в Западной Азии. Азербайджанцы генетически родственны тюрко- и ираноязычному населению Западной Азии, но отличаются от туркмен. А иранские азербайджанцы имеют генетическое сходство со своими ираноязычными соседями, в то время как представители этнографической группы азербайджанцев Дагестана - терекеме демонстрируют генетическое сходство с кумыками.
1/3
В целом опубликованные данные свидетельствуют о том, что 18% генофонда азербайджанцев отражают средневековые миграции из Центральной Азии и 6% связаны с миграциями из Восточной Азии.
А полногеномные данные азербайджанцев Северо-Западного Ирана рассмотренные в работе, посвященной следам тюркской экспансии, указали на 5%-й вклад восточноевразийской предковой компоненты, что можно объяснить распространением тюркских языков на запад согласно модели «доминирования элит» (это когда происходит смена языка без значительного изменения генофонда).
При этом генетических данных из Азербайджана было очень мало. А ведь именно эта территория является значимой для выявления многовекового влияния Персии на регион, в том числе и с точки зрения генетики. При этом интересно: распространялось ли это генетическое влияние только на южные территории или на весь Каспийский регион? Кроме того, данные из восточных регионов Кавказа и Ирана могут также прояснить и генетическую историю ряда малочисленных народов Северного Кавказа.
В более изученных популяциях Дагестана обнаружена значимая корреляция между частотами гаплогрупп, их географией и языком (о чем свидетельствуют лексикостатистические данные).
При этом несмотря на сильное межпопуляционное разнообразие коренных народов Дагестана была подтверждена гипотеза о происхождении всех нахско-дагестаноязычных популяций от одной прапопуляции около 6000-6500 лет назад. Которая в свою очередь восходит к единой послеледниковой предковой популяции, которая претерпела изменения из-за смены ландшафта, генетического дрейфа и эффекта основателя. В отличие от горных народов Дагестана, его равнинные популяции обладают большим разнообразием Y-хромосом, что можно объяснить их контактами с населением Центральной Азии. Дидойцы или цезы, представляющие аваро-андо-цезскую ветвь нахско-дагестанской языковой семьи, демонстрируют более скромное генетическое разнообразие из-за преобладания гаплогруппы J1 и значительного эффекта основателя.
Две работы этого года, опубликованные с интервалом в несколько месяцев, касаются аутосомного генофонда народов Восточного Кавказа и изменчивости Y-хромосомы в тех же популяциях. Такой систематический анализ генофондов популяций, населяющих исторически связанные территории восточной части Кавказа позволяет увидеть наиболее объемную и объективную картину изменчивости генофонда популяций этой физико-географической области. А результаты этих исследований могут быть использованы для моделирования расселения ранних людей по Кавказу, Западной Азии и Евразии, используя обновленные и расширенные данные ДНК.
Всего в работах было проанализировано 2216 человек из 22 этнических групп Восточного Кавказа и сопредельных регионов, представляющих 3 языковые семьи: нахско-дагестанскую (аваро-андо-цезская, лакская, лезгинская и нахская ветви), алтайскую (тюркская ветвь) и индоевропейскую (иранская группа).
В исследованиях основное внимание уделялось Восточному Кавказу, генетические данные 1329 человек, представляющих коренные народы Дагестана и Азербайджана, были проанализированы более подробно, чем популяции степей Северного Каспия, носители нахских языков и ираноязычные народы Кавказа, которые вошли в группы сравнения и были представлены объединенными выборками без дальнейшего разделения на более мелкие подгруппы.
Нахско-дагестанская языковая семья была представлена 14 этническими группами: аварцами и тиндинцами (аваро-андийская языковая подветвь), цезами и гинухцами (цезская подветвь), даргинцами, кайтагцами и кубачинцами (даргинская ветвь), лакцами (лакская ветвь), лезгинами, рутулами, табасаранами и цахурами (лезгинская ветвь нахско-дагестанской семьи), а также чеченцами и ингушами (нахская ветвь). Тюркоязычные народы были представлены населением Восточного Кавказа (караногайцами, кумыками, терекеме Дагестана и азербайджанцами Азербайджана) и западной части Евразийской степи (ставропольскими ногайцами, астраханскими татарами, ногайцами и ставропольскими туркменами или трухменами). Ираноязычное население Кавказа было представлено татами, горскими евреями, езидами, курдами и талышами.
А в исследовании аутосомного генофонда 22 этнические группы Восточного Кавказа представлены данными о 243 геномах по широкогеномным панелям SNP-маркеров, в дополнение к 43 ранее опубликованным, которые были изучены в основном по узким панелям маркеров.
Более половины изученных образцов опубликованы в новых работах впервые.
Гаплогруппы Y-хромосомы в популяциях Восточного Кавказа и прилегающих регионов
Географическая изменчивость проанализированных гаплогрупп Y-хромосомы на Восточном Кавказе неодинакова для представителей разных языковых семей: она более выражена для нахско-дагестанских языковых групп, чем для тюркоязычного и ираноязычного населения.
В целом, авторы выделили 5 основных гаплогрупп с различными географическими моделями распределения: J1, J2, R1a, R1b и N. Их разнообразие продемонстрировано на картах частотного распределения.
J1
Для гаплогруппы J1 характерна очаговая структура распространения, в Дагестане её частота снижается во всех направлениях от центра к периферии.
Возраст гаплогруппы J1 составляет от 9000 до 24000 лет и, как было показано в предыдущих исследованиях, она возникла в Западной Азии, где ее ветвь J1-P58 является наиболее распространенной и достигает пика частоты у арабов Ближнего Востока. Однако линия J1-P58 крайне редка для популяций Восточного Кавказа. Хотя в целом на долю линий гаплогруппы J1-M267(×P58) приходится более 50% генофонда дагестаноязычных популяций. За исключением цахуров (44%) и табасаран (13%). А максимума частота J1-M267(×P58) достигает у кубачинцев (98%), ареал обитания которых ограничен селом Кубачи. J1-M267(×P58) также преобладает и в других популяциях Дагестана (от 59 до 92%) и часто встречается среди чеченцев (17-25%). Однако её частота резко снижается по мере удаления от носителей нахско-дагестанских языков. При этом высокая частота гаплогруппы J1-M267(×P58), наблюдаемая у кумыков, может указывать на значительный генетический вклад нахско-дагестаноязычных популяций, поскольку эта гаплогруппа обычно не встречается в других тюрко- и ираноязычных популяциях Восточного Кавказа.
Высокая доля гаплогруппы J1-M267(×P58) в популяциях Восточного Кавказа может объясняться не только действием дрейфа генов, но и наличием иных – кроме J1-P58 - ветвей в данном регионе. Поскольку ранние носители гаплогруппы J1 появились на Кавказе еще в палеолите (~13 тыс. лет назад; охотники-собиратели на территории современной Грузии), а возникновение земледельческих сообществ в неолите привело к росту населения и появлению новых гаплогрупп, возможно, несколько местных линий возникли в пределах J1-M267(×P58). К этому вопросу ещё вернёмся.
J2
Гаплогруппа J2-M172(×M67, M12) также имеет очаговую структуру, но ее пиковые частоты приходятся на юг региона: совокупный вклад и разнообразие ветвей J2 больше у азербайджанцев и ираноязычного населения Кавказа. Она редко встречается в популяциях Дагестана, но ее ветвь J2-M67(×M92) встречается с частотой 67% у их соседей — чеченцев и ингушей, носителей нахских языков. Происхождение гаплогруппы J2 прослеживается до Западной Азии, где наблюдается наибольшее разнообразие её ветвей (J2a-M92 и J2a-M530). Её частота достигает максимума в ираноязычных популяциях.
В Западной Азии основными носителями гаплогруппы J2 являются турки, азербайджанцы, курды и персы (20-35%). Наиболее распространенной является ветвь J2-M172 (×M67, M12) встречающаяся с частотой 15 и 20% среди тюркоязычных и ираноязычных популяций соответственно. А на Восточном Кавказе её частота составляет 11% у тюркоязычных и 17% у ираноязычных народов. Таким образом, частоты гаплогруппы J2 и ее ветви J2-M172(×M67, M12) среди азербайджанцев Дагестана - терекеме и азербайджанцев Азербайджана близки к частотам, наблюдаемым не только у ираноязычных народов Кавказа, но и у населения Западной Азии.
R1a и R1b
В целом, гаплогруппы R1a и R1b выявляют связь между степными популяциями на севере и группами, расселившимися вдоль Каспийского побережья на юге. Их совместный вклад наиболее значителен для тюркоязычных народов Дагестана (24-40%), азербайджанцев Азербайджана (30%) и ираноязычного населения Кавказа (37%). Их частота плавно снижается в нахско-дагестанских популяциях Дагестана, за исключением табасаран.
R1b
Гаплогруппа R1b представлена 2 субкладами: R1b-M269 и R1b-M73, из которых субклада M269 наиболее распространена. Её максимальная частота наблюдается у табасаран (64%), однако у большинства других носителей нахско-дагестанских языков не превышает 15%. R1b-M269 встречается у ираноязычных популяций Кавказа с частотой в 26%, азербайджанцев (23%) и кумыков (19%). Помимо этого, эта линия встречается с высокой частотой на юге региона среди азербайджанских талышей (48%).
В целом во всей Западной Азии вклад R1b значительно выше для тюркоязычного населения (20% для азербайджанцев и турок), чем для ираноязычного (5%). Схожие частоты R1b-M269 среди ираноязычного населения Кавказа и тюркоязычных народов Западной Азии могут указывать на генетические взаимодействия между их предковыми популяциями. R1b-M73 встречается с заметной частотой среди караногайцев (13%), популяций северокаспийских степей и тюркоязычных популяций Западного Кавказа (карачаевцев и балкарцев).
R1a
Гаплогруппа R1a представлена в основном ветвью R1a-M198(×M458), которая очень распространена среди карапапахов (19%) и караногайцев (18%), но встречается с меньшей частотой в других популяциях Восточного Кавказа, проанализированных в этой работе (2-11%), за исключением цезов, кубачинцев и табасаран. Гаплогруппа R1a широко распространена в Евразии: за исключением ветви R1a-M458, которая типична для популяций Восточной Европы и редко встречается в популяциях Восточного Кавказа. R1a включает в себя удивительное разнообразие линий, охватывающих обширную географию от Индии до Скандинавии.
N
Гаплогруппа N достигает самых высоких частот в степной популяции караногайцев на севере Дагестана (22%) и азербайджанских карапапахов (10%). Однако у них разные линии: у караногайцев субклады N3a5a-F4205 и N3a2-M2118, распространенные в Сибири и Центральной Азии, а также N-LLY22g (×M178, Y3205); а у карапапахов N2-Y3205 и N3a1-B211, которые являются типичными для популяций Уральского региона и Западной Сибири.
Основные закономерности географической изменчивости
Карты генетических расстояний Нея, которые объединяют данные по всем гаплогруппам, показывают степень сходства и различия между изучаемыми популяциями, а также дают общую характеристику генофонда Y-хромосомы для каждой популяции и выделяют регионы с наибольшим генетическим сходством.
В целом генофонд Восточного Кавказа представлен тремя основными паттернами: дагестанским, иранским и степным.
Дагестанский паттерн
Карты генетических расстояний между говорящими на языках, входящих в нахско-дагестанскую языковую семью, имеют общую закономерность, единственным исключением является табасараны.
Этот дагестанский паттерн характеризуется близкими генетическими расстояниями между популяциями, встречается только в Дагестане и не коррелирует с языком. Похожая, но менее выраженная картина у тюркоязычных кумыков. Дагестанский паттерн показывает глубокую связь между генофондами популяций Дагестана с точки зрения гаплогрупп Y-хромосомы. Эта закономерность в значительной степени определяется присутствием гаплогруппы J1-M267(×P58). В рамках данного исследования наблюдается генетическое сходство между популяциями Дагестана и Северного Ирана, которое определяется вкладом гаплогрупп J1 и J2 и может отражать самые ранние волны миграции на Восточный Кавказ.
В целом дагестанский паттерн указывает на связь с древним автохтонным кавказским населением породившем множество генофондов благодаря мощному генетическому дрейфу, которые до сих пор сохраняют свое единство.
Иранский паттерн
Существует генетическое сходство между всеми азербайджанскими и ираноязычными популяциями Кавказа (татами, талышами, езидами, курдами). Вклад гаплогрупп J2-M172 (×M67, M12) и R1b-M269 в этот паттерн является наибольшим, что можно увидеть, сравнив карты их распределения и генетических расстояний. Эта закономерность проявляется на карте генетических расстояний от табасаран, которые представляют лезгинскую ветвь нахско-дагестанских языков, хотя для табасаран эта закономерность менее выражена.
Иранский паттерн связан с древним иранским населением, пришедшим из Мидии, и более ранними волнами миграции в 3-2 тысячелетиях до нашей эры.
Степной паттерн
Карты генетических расстояний также демонстрируют значительные различия между тюркоязычными популяциями. Так, генофонд кумыков близок к генофонду кавказоязычного населения Дагестана, а генофонд азербайджанцев схож с генофондом ираноязычных народов Кавказа. Единственная закономерность обнаружена у караногайцев и отражает весьма отчетливую связь с генофондами евразийской степи. На значительно более слабом уровне такая закономерность есть и у азербайджанских карапапахов. Степной паттерн отражает одну из последних миграционных волн тюркского населения и является более поздним с точки зрения своего происхождения и вклада в генофонд Восточного Кавказа.
Восточный Кавказ в многомерном генетическом пространстве
Картографический анализ генетических расстояний выявляет на одной карте области генетического сходства одной популяции. В свою очередь, визуальное представление всей попарной матрицы генетических расстояний Нея посредством многомерного шкалирования показывает положение всех изученных популяций. На графике наблюдается три кластера — дагестанский, иранский и степной, которые отражают географические, а не лингвистические закономерности.
1/2
Северо-восточный вектор влияния Евразийской степи проявляется в степном кластере. Юго-восточный вектор отражает влияние Западной Азии и формирует произвольный иранский кластер. Вектор автохтонного кавказского населения проявляется в дагестанском кластере, объединяющем все дагестаноязычные народы. При этом между иранским и степным кластерами генетическое расстояние меньше, чем между иранским и дагестанским. А максимальное расстояние между дагестанским и степным кластерами. Аналогичные закономерности наблюдаются и при анализе главных компонент.
Степной кластер образован караногайцами Дагестана и объединенными популяциями тюркоязычных народов, населяющих север Прикаспийской степи (туркмены, ногайцы и астраханские татары). Кластер характеризуется наибольшей неоднородностью и отражает миграции внутри евразийских степей, в том числе средневековые миграционные волны, повлиявшие на генофонды ногайцев и астраханских татар, новейшую миграцию туркмен из Прикаспийского региона в XVII в. и др. Прошлые генетические взаимодействия между популяциями Восточного Кавказа и евразийских степей проявляются только на северной границе региона в генофонде дагестанских караногайцев и гораздо слабее у кумыков и карапапахов.
Иранский кластер включает представителей трех языковых семей: тюркоязычных азербайджанцев из Дагестана и Азербайджана, ираноязычного населения Кавказа и табасаран, говорящих на языке лезгинской группы нахско-дагестанской семьи. Этот кластер намного плотнее, чем степной, но менее объединенный, чем дагестанский. Присутствие табасаран в этом кластере может отражать наследие Табасаранского майсумства (государственного образования на юге Дагестана), где они жили рядом с другими популяциями кластера, или, альтернативно, может указывать на влияние средневековых миграций из провинции Табаристан на севере Ирана. Иранский кластер отражает взаимодействие между местным населением и населением Западной Азии и, следовательно, может быть назван иранским. Народы Восточного Кавказа и Западной Азии поддерживали свои контакты на протяжении длительных периодов истории; их контакты происходили на обширной территории, простирающейся за пределы Восточного Кавказа, поэтому необходимы дальнейшие исследования для изучения источников и истории этого компонента.
Дагестанский кластер включает в себя все кавказоязычные популяции Дагестана, за исключением табасаран, и тюркоязычных кумыков Дагестана, которые географически близки друг к другу. Распределение популяций внутри кластера отдаленно напоминает их географию: проживающие севернее даргинцы, лакцы и цезы, занимают условно “северо-западную” часть графика. Лезгинские популяции, расселенные дальше к югу, занимают “юго-восточную” часть графика, то есть противоположную часть скопления. Положение кумыков на границе кластера представляет собой основной пласт их генофонда, связывающий их с кавказоязычным населением и предполагающий генетические «заимствования» у других тюркоязычных популяций. Нахские народы (чеченцы и ингуши) расположились выше границ кластера.
Географически плотный дагестанский компонент генофонда Восточного Кавказа, встречающийся в большинстве дагестанских популяций, в значительной степени определяется высокой частотой гаплогруппы J1-M267 (×P58) поэтому рассмотрим её структуру более подробно.
Филогеография гаплогруппы J1-M267(×P58)
Ветвь J1-Y3495, возрастом 6,4 ± 0,6 тыс. лет, является наиболее распространенной в регионе.
До сих пор субклада J1-Y3495 не была подробно описана в литературе, но ее предковая линия J1-Z18375 (или филогенетический эквивалент Z1841) хорошо известна и авторы сосредоточились на ней. Для J1-Z1841 время до ближайшего общего предка по разным расчётам составляет от 8 до 6,5 тыс. лет. Эта линия обнаружена у одного из трех представителей куро-араксской культуры (село Великент Дербентского района Дагестана), относящейся к эпохе бронзы (~5000 лет назад).
Это натолкнуло авторов работы на предположение о географическом происхождении J1-Z1841 на Кавказе или в его окрестностях. Её сублиния J1-Z1842 была обнаружена у человека на востоке Анатолии или Малой Азии, жившего ~5000 лет назад. Тот факт, что линия J1-Z1841/Z18375 была распространена в Малой Азии и Леванте около 3000 лет назад, предполагает контакты между носителями хуррито-урартских и нахско-дагестанских языков.
Единственной линией, представляющей гаплогруппу J1-Z1841 в популяциях Восточного Кавказа, является J1-Y3495. Авторам удалось идентифицировать 17 полиморфных линий этой гаплогруппы.
Карты показывают, что линии J1-ZS3114 и J1-CTS1460, а также J1-Y3495 (×ZS3114, CTS1460), возрастом от 6400 до 5900 лет, встречаются почти повсеместно в регионе: J1-ZS3114 более распространена среди носителей даргинской, лакской и лезгинской ветвей нахско-дагестанской семьи языков; J1-CTS1460 типична для носителей языков аваро-андо-цезской ветви той же семьи; J1-Y3495(×ZS3114, CTS1460) широко распространена среди носителей даргинской ветви и несколько менее часто встречается у носителей лезгинской ветви нахско-дагестанской семьи.
География этих трех линий пересекается с заметной частотой (более 20%) в Дагестане, что позволяет предположить, что линия J1-Y3495, возможно, появилась на его территории.
Небольшая разница, около 500 лет, в средних датах происхождения между гаплогруппой J1-Y3495 и двумя ее ветвями J1-ZS3114 и J1-CTS1460, указывает на рост населения в регионе в раннем бронзовом веке (~6 тыс. лет назад), во время появления новых методов обработки металла, экономических и социальных изменений, включающих социальное неравенство и племенные объединения, а также торговых отношений с населением Юго-Восточной Европы и Западной Азии.
Лингвистические данные свидетельствуют о том, что раскол нахско-дагестанского праязыка произошел в конце 3-го тысячелетия до н.э. (~5 тыс. лет назад), что согласуется с новыми генетическими данными: более древние линии, возрастом ~6000 лет, встречаются почти во всех дагестанских популяциях, проанализированных в этой работе, и у некоторых нахских народов, хотя пики частот для J1-CTS1460 и J1-ZS3114 наблюдаются в ряде различных языковых групп.
Таким образом, основная часть генофонда населения Дагестана по отцовской линии (носителей гаплогруппы J1-M267(×P58)) может быть прослежена до предковой линии J1-Y3495. Авторы предполагают, что в позднем медном веке и раннем бронзовом веке (~6,5 тыс. лет назад) носители этой линии расселились по центральной части горного Дагестана, где география основных субклад J1-Y3495 пересекается.
При этом носители этой линии были частью сообщества, говорившего на пра-нахско-дагестанском языке.
Похожий вывод делали и другие учёные в работах с более скромными техническими и методологическими возможностями. Наличие одной основной гаплогруппы в генофонде Дагестана может свидетельствовать о сокращении древней популяции в более ранние периоды, когда другие линии, сосуществовавшие с J1-Y3495, вымерли из-за эффекта бутылочного горлышка. Т.е. могло иметь место сокращение численности популяции с последующим её восстановлением, но с меньшим генетическим разнообразием.
Эффект бутылочного горлышка с последующим ростом популяции, но с меньшим генетическим разнообразием.
Географические особенности распространения наиболее часто встречающихся вариантов линии J1-CTS1460 (Y6916 (×ZS2872), Y61473, ZS2910 (×ZS2878, ZS7652) и ZS2878) показывает, что ее географический центр находится в районах, занятых людьми, говорящими на языках аваро-андо-цезской ветви нахско-дагестанских языков, и распространяется на некоторые районы южнее. Датировка линий около 4-5 тыс. лет назад предполагает непрерывный рост популяций после их разделения.
В первой части обзора, мы рассмотрели отцовские линии, однако не только мужчины участвуют в формировании всего генофонда популяций, как уже отмечалось ранее, не все линии гаплогрупп Y-хромосомы дошли до наших дней, однако в аутосомном генофонде остаются следы всех его родоначальников в той или иной мере. Поэтому во второй части рассмотрим общегеномные данные в этих же популяциях Восточного Кавказа.
Источник:
Agdzhoyan, A.; Iskandarov, N.; Ponomarev, G.; Pylev, V.; Koshel, S.; Salaev, V.; Pocheshkhova, E.; Kagazezheva, Z.; Balanovska, E. Origins of East Caucasus Gene Pool: Contributions of Autochthonous Bronze Age Populations and Migrations from West Asia Estimated from Y-Chromosome Data. Genes 2023, 14, 1780. https://doi.org/10.3390/genes14091780
Приветствую! Сегодня речь пойдёт о научной работе, авторы которой рассматривают фундаментальный вопрос о происхождении и особенностях миграций носителей фатьяновской и средневолжской абашевской культур. Цель работы заключалась в том, чтобы на основании новых лабораторных данных о древней ДНК рассмотреть гипотезы о пришлом происхождении палеопопуляций фатьяновской и средневолжской абашевской археологических культур на Русской равнине.
В исследовании были проанализированы образцы 25 человек, относящихся к фатьяновской культуре (20 образцов было получено от захороненных в пределах Волосово-Даниловского могильника, два из раскопок в Никульцино и по одному из Воронково, а также Голузинского и Горицкого могильников). Стоит отметить, что Волосово-Даниловский могильник — наиболее репрезентативный погребальный комплекс фатьяновской археологической культуры.
Помимо этого, впервые был проведен палеогенетический анализ 14 человек, относящихся к абашевской культуре (тринадцати человек из Пепкинского кургана и одного из Старшего Никитинского могильника). Пепкинский курган, открытый у деревни Пепкино, представляет собой коллективное захоронение группы молодых мужчин, не менее 27, средневолжской абашевской археологической культуры, погибших в бою.
Кстати, для исследования авторы работы намеренно отбирали представителей мужского пола разного возраста, так как для них возможен анализ однородительских маркеров как по мужской, так и по женской линиям.
Здесь сразу хотелось бы выразить благодарность авторам работы за их вклад в наше понимание прошлого.
Введение
Обнаружение в геномах европейских групп компонента восточных охотников и собирателей, соотносимого с генетическим наследием населения ямной археологической культуры, в своё время распространенной на обширных территориях евразийских степей, стало одним из наиболее спорных вопросов в российской археологии.
И хотя между распространением культуры и генов нельзя ставить знак равенства, генетический вклад от носителей ямной культуры зафиксирован во многих регионах Европы. Наличие этого степного генетического сигнала отличает представителей культур шнуровой керамики и колоколовидных кубков, а также более поздних жителей Европы эпохи бронзы от ранних неолитических земледельцев этого региона. При этом на территории европейской части современной России подобные генетические исследования до недавнего времени отсутствовали, пока в 2021 году не была опубликована работа, обзор которой также есть на канале, под названием: Генетическая история населения запада России при переходе от каменного века к бронзовому. В этом, более раннем, исследовании авторы пришли к выводу о зарождении носителей фатьяновской культуры благодаря миграции смешанного населения, потомков носителей ямной культуры и европейских ранних земледельцев.
Тогда авторы сообщали, что в моделях только две популяции, а именно из культуры шаровидных амфор Украины и Польши, а также трипольской культуры Украины, оказались потенциальными источниками для смешения со степными скотоводами с генетическими профилями как у представителей ямной культуры Самары или Калмыкии, для образования генетической структуры популяции фатьяновцев. Также была отмечена близость этих людей с представителями культуры шнуровой керамики.
На основании данных археологии в последние годы уже высказывались предположения, что на территории Восточно-Европейской равнины в конце III тыс. до н. э. происходили масштабные миграции, с первоначальным импульсом из центральноевропейского региона. В пользу этого довода говорит схожесть керамического комплекса средневолжской абашевской культуры, следы которой выявлены непосредственно в Поволжье на территориях Брянской, Московской и Ярославской областей, с керамикой поздней фазы культуры колоколовидных кубков в Южной Германии.
Результаты
Анализ Y-хромосомы
По результатам микросателлитных исследований и анализа Y-хромосомы по методу секвенирования нового поколения (NGS) для образцов, относящихся к фатьяновской культуре, получена единственная гаплогруппа R1a (Z93).
Вероятно, такая однородность гаплогрупп Y-хромосомы связана с эффектами основателя и дрейфа генов в изолированной древней группе неместного происхождения.
Эффект основателя — вариант генетического дрейфа, при котором снижение и смещение генетического разнообразия происходит из-за заселения малым количеством представителей вида новой географической территории.
Дрейф генов — Изменение частот аллелей и генотипов в популяции, имеющее случайный характер. Такие случайные отклонения в частотах аллелей происходят из поколения в поколение, особенно при небольшом эффективном размере популяции.
На данный момент линии гаплогруппы R1a1a1b2 Z93 с заметной частотой встречаются в Южной Сибири среди алтайцев и хакасов (>30 %), также сравнительно небольшие частоты встречаются среди татар Поволжья (5 %), в Кыргызстане (6 %) и во всех популяциях Ирана (1—8 %).
Среди археологических культур указанная гаплогруппа обнаружена, в частности, в сагсайской культуре XV—XII вв. до н. э. Субветвь этой гаплогруппы R1a1a1b2a Z94 представлена рядом образцов, относящихся к археологическим культурам средней бронзы центральной и западной части степного коридора Евразии — андроновской, полтавкинской, синташтинской и также сагсайской (XX—XV вв. до н. э.).
А вот для образцов абашевской культуры полученные результаты менее однородны. Для большинства образцов из Пепкинского кургана определена гаплогруппа R1a (Z93>Z94), однако у троих мужчин с этого участка была гаплогруппа R1b (Z2103).
Примечательно, что для абашевца захороненного на значительном удалении от Пепкинского кургана в Старшем Никитинском могильнике, был получен достаточно полный гаплотип, также предсказывающий принадлежность к той же гаплогруппе R1b (Z2103).
Указанная гаплогруппа была выявлена у мужчин, относящихся к ямной, афанасьевской, полтавкинской, катакомбной и окуневской культурам III—II тыс. до н. э.
Микросателлитные исследования в палеогенетике являются важным инструментом, особенно при исследовании групповых захоронений, поскольку позволяют выявить близость с точки зрения происхождения по мужской линии, а иногда и прямого родства, когда гаплотипы по всем считанным STR локусам совпадают, при этом результат тем точнее, чем выше число считанных локусов. Такие совпадения были найдены как для фатьяновцев погребенных в Волосово-Даниловском могильнике, так и для абашевцев из Пепкинского кургана, а также Старшего Никитинского могильника.
Что указывает на возможное близкое родство по отцовской линии людей с совпадающими STR локусами. А у людей, которые продемонстрировали минимальные отличия в значениях аллелей в одном или двух локусах, общий предок по отцовской линии мог жить несколькими поколениями ранее.
Анализ митохондриальной ДНК
Материнские линии фатьяновцев были более разнообразны, чем отцовские. При этом основной гаплогруппой мтДНК была U с сублиниями — U5a1b, U5a1d2a и U5a1a1. А среди редких, так называемых минорных линий, определены H6a1a и I1a1
В большинстве линии гаплогрупп совпадают с задокументированными и в работе 2021 года, но с некоторыми дополнениями, как линии U5a1d2a и U4a1 у фатьяновцев из Волосово-Даниловского могильника. Однако в предыдущем исследовании линия U4a1 определена у одного из охотников-собирателей культуры Веретье эпохи мезолита, захороненного в могильнике Песчаница 1 на юго-западе Архангельской области. А ещё ранее эта линия была обнаружена у восточных и скандинавских охотников-собирателей. В частности, у охотников и собирателей эпохи неолита Большого Оленьего острова в Кольском заливе.
А что касается хронологически более поздних представителей абашевской культуры, то часть их линий, таких как U5a2b2, H41, ранее были обнаружены и у фатьяновцев. Однако неожиданностью для авторов стало наличие гаплогрупп J2b1a2a и K1b1a1+199 у абашевцев из Пепкинского кургана, ведь распространение этих линий связывают с неолитическими земледельцами. При этом пары с этими линиями имеют полностью совпадающий набор мутаций, что указывает на их родство по материнской линии.
Анализ аутосомных маркеров
Сравнительный анализ аутосомных генетических вариантов с использованием метода понижения размерности проводился для 19 образцов в сочетании с ранее опубликованными данными 75 человек. По результатам анализа большая часть исследованных в новой работе образцов фатьяновской и абашевской культур объединились между собой в одном кластере.
Туда же вошли некоторые фатьяновцы из предыдущей работы. Неожиданный результат был получен для двух фатьяновцев из Волосово-Даниловского могильника, которые оказались ближе к скоплению образцов из контекста различных археологических культур эпохи бронзы, а именно из культур шнуровой керамики, колоколовидных кубков, афанасьевской и синташтинской. Что указывает на генетическую близость жителей эпохи бронзы, несмотря на их разную хронологию и географию.
Обсуждение и выводы
В новой работе у представителей фатьяновской культуры не обнаружено других отцовских линий, кроме R1a (Z93), что, вероятно, связано с эффектом основателя (когда небольшая группа отделяется от первоначальной популяции и формирует новую) и с дрейфом генов (когда одна линия сохранилась из-за случайных событий).
Это подтверждает миграцию группы представителей культуры шнуровой керамики из Центральной Европы и отсутствие связей с представителями местной волосовской культуры, что также упоминалась и в предыдущем исследовании, хотя его авторы использовали другие образцы и методы. Примечательно, что ранее, по результатам анализов травм левой руки в области локтевого сустава и кисти у мужчин из Волосово-Даниловского могильника, авторы исследования, опубликованного в 2022 году, пришли к выводу о схожей тактике боя у фатьяновцев Верхней Волги и шнуровиков Германии и Польши. Что служит независимым индикатором их принадлежности к единой культурной общности.
Помимо этого, совпадение митохондриальных гаплогрупп в обоих упомянутых генетических исследованиях также подтверждает гипотезу о неместном происхождении носителей фатьяновской культуры. При этом однообразный погребальный обряд фатьяновцев, различный для мужчин, женщин и детей, а также их низкое генетическое разнообразие, добавляют аргументов в пользу существования строгих правил внутри общества, которые можно объяснить его сплочённостью в связи с миграцией в иную среду. А учитывая некоторую близость по материнской линии захороненных в Волосово-Даниловском могильнике, с преобладанием ветвей гаплогруппы U, можно сделать вывод, что фатьяновцы переселялись семьями.
Датировка образцов из Волосово-Даниловского могильника указывает на его использование в течение более трёхсот лет, минимум с середины третьего тысячелетия до н. э. (интервал калиброванных дат: 2563-2037 гг. до н. э.).
При этом Волосово-Даниловский могильник всеми исследователями рассматривался как наиболее поздний.
А спустя несколько сотен лет после первого появления фатьяновцев возникла абашевская археологическая культура, в лесостепных районах среднего Поволжья и верхнего Дона, предположительно, основанная другими мигрантами. При этом Пепкинский курган, на котором были захоронены мужчины, погибшие в одном сражении, на данный момент является главным археологическим памятником средневолжской абашевской культуры. На первый взгляд это могли быть воины одного племени, потому как они захоронены в одной могиле и по схожему обряду, между 2130-1950 гг. до н. э.
Однако анализ древней ДНК выявил как минимум две группы разного происхождения. Экспертиза бронзовых изделий из Пепкинского кургана и Старшего Никитинского могильников показала, что по составу металла оба памятника относятся к единому этапу средневолжской абашевской культуры. А мышьяк и серебро в составе их металла, могут указывать на связь с рудными источниками Южного Урала. При этом для образца из Старшего Никитинского могильника была получена несколько более поздняя дата чем для Пепкинского кургана, отличающаяся на время жизни одного поколения абашевцев (3557 ± 31 BP).
Примечательно, что в Пепкинском кургане, один из абашевцев, погребенный с набором предметов кузнеца-бронзолитейщика с гаплогруппой Y-хромосомы R1b (Z2103), по данным изотопного анализа провел свои юные годы в геохимических условиях, которые соответствовали степным районам Южного Урала.
К тому же, известно о происхождении металла некоторых изделий, найденных в Пепкинском кургане, из Таш-Казганского месторождения меди.
При этом по соотношениям изотопов стронция в зубной и костной ткани другой группы мужчин из Пепкинского кургана, все они оказались «неместными» для территории современной республики Марий Эл, где они были похоронены. Примерно до 10 лет они жили в регионе соответствующему западному ареалу проживания абашевцев в бассейне Оки. Похожие значения характерны и для захороненных в пределах Старшего Никитинского могильника. Интересно, что абашевцы с гаплогруппой R1b(Z2103) были более мобильны, но при этом связаны достаточно тесным родством. Можно предположить, что именно эта группа в составе абашевского населения была тесно связана с металлургическим производством. Однако другая, более многочисленная часть мужчин из Пепкинского кургана с гаплогруппой R1a (Z93>Z94), очевидно, имела иное происхождение. Отцовские линии и аутосомные маркеры сближают их с фатьяновцами, прежде всего, из Волосово-Даниловского могильника. Однако неясно, связано ли происхождение абашевцев с фатьяновцами, что в целом не подтверждается археологически, или же с новой миграцией из Европы спустя несколько веков после миграции предков фатьяновцев. И хотя митохондриальные линии поддерживают обе гипотезы, отдельные линии обнаружение у некоторых людей из Пепкинского кургана, такие как J2b1a2a и K1b1a1+199, скорее, свидетельствуют о недавней миграции из Европы, где генетический след неолитических земледельцев был очень заметен.
Ранее была предложена гипотеза общей энеолитической подосновы населения абашевской культуры и культур шнуровой керамики.
А по черепам из Пепкинского кургана антропологами была отмечена высокая изменчивость у абашевских мужчин.
Графические реконструкции мужчин погребенных в Пепкинском кургане (Реконструкции Г. В. Лебединской).
Но, в целом, метод главных компонент выявил сближение выборки мужчин, погребенных в Пепкинском кургане, с южноевропеоидными формами, ранее отнесенными к грацильным представителям средиземноморской расы. Что больше свидетельствует в пользу недавней миграции из Европы.
На графике с использованием метода понижения размерности (UMAP) люди из Волосово-Даниловского могильника сближаются со второй, более многочисленной группой из Пепкинского кургана, что может указывать на преемственность. Однако абашевец из погребения 7 Пепкинского кургана близок и к мужчине унетицкой культуры из Польши, а также к представителю позднего неолита из Марбьерга в Дании, которые жили примерно в то же время, что и он. А это уже аргумент в пользу недавней миграции.
У носителей фатьяновской культуры, на протяжении всего периода существования Волосово-Даниловского могильника, наблюдается достаточно стандартный погребальный обряд, который роднит их с культурами шнуровой керамики Европы, чего нельзя сказать для набора инвентаря поселений фатьяновцев. Но обряд свидетельствует о формировании могильника представителями пришлого населения, несущего традиции европейских групп шнуровой керамики.
А вот вопрос о происхождении представителей средневолжской абашевской культуры решается не столь однозначно. В ее составе присутствуют две составляющие — с гаплогруппами R1a (Z93>Z94) и R1b (Z2103).
Для носителей линии R1a (Z93>Z94) типично захоронение в могиле рядом близких родственников по материнской линии (причем здесь выделяются пары с европейскими «раннеземледельческими» митохондриальными гаплогруппами: как J2b1a2a из погребений 20, 21 и K1b1a1+199 из погребений 16,17).
В то время как захороненных рядом мужчин с гаплогруппой Y-хромосомы R1b (Z2103) сближает родство по отцовской линии. Это указывает на то, что система родства в группах разного происхождения отличалась.
Примечательно, что абашевцы из Пепкинского кургана с гаплогруппой R1a (Z93>Z94), в том числе, связанные тесным родством, всегда имели с собой костяные поделки и украшения, в отличие от носителей линии R1b (Z2103), у отдельных носителей которой прослеживаются связи со степными районами Южного Урала.
Однако решение вопроса о происхождении двух составляющих абашевского населения лежит в плоскости будущих исследований.
Источник: А. В. Энговатова, И. Э. Альборова, Х. Х. Мустафин, В. Ю. Луньков, Ю. В. Лунькова (Москва, Россия), А. А. Канапин, А. А. Самсонова (Санкт-Петербург, Россия), М. Б. Медникова (Москва, Россия) Древняя ДНК носителей фатьяновской и абашевской культур (к вопросу о миграциях населения эпохи бронзы в лесной полосе на Русской равнине) doi.org/10.55086/sp232207228
Взять с собой побольше вкусняшек, запасное колесо и знак аварийной остановки. А что сделать еще — посмотрите в нашем чек-листе. Бонусом — маршруты для отдыха, которые можно проехать даже в плохую погоду.
Довольно любопытный вопрос прозвучал под статей @Aurinel, и реакция на ответ показала наличие заблуждений. Что ж, попробую расписать подробней, как сочетаются этнос, генетика и язык.
Собственно, сам вопрос:
Чтобы полностью запутать, необходимо расширить этот вопрос до наличия связи между этносом, генетикой и языком.
Спойлер для ЛЛ: это пересекающиеся множества
Распутывать связь необходимо от самого понятия этноса.
Этнос
Различные исследователи имеют разнообразные подходы к тому, что же считать этносом и как его определять. В принципе, все подходы совпадают лишь в одном: этнос - это некая совокупность людей, которую можно отличить от другой совокупности людей. Часто это понятие смешивают с понятиями народ, народность, национальность, этничность.
В этом ряду сильно выбивается в сторону только термин "национальность", т.к. он изначально относится к государственной принадлежности, но в русском языке получил дополнительное значение.
Обычно, когда исследователи говорят об этносе, они упоминают признаки, позволяющие отделить один этнос от другого. Чаще всего выделяются следующие: происхождение, территория проживания, культура, внешний вид, язык, самосознание. Пройдемся по каждому признаку.
Логично, что этносы зарождаются где-то в глубине истории. Обычно мы можем даже сказать, какой этнос примерно где и когда родился. Так, к примеру, про русский этнос говорят, что он был рожден во времена монгольского ига, когда на Куликовскую битву пришли люди разных княжеств, а вышел оттуда русский народ. Этнос может родиться как за счет выделения из какой-нибудь совокупности людей, так и за счет смешения.
Отсюда логичным продолжением является, что у этноса есть некая территория проживания, ограниченная внешними факторами. На этом шаге этнос совпадает с понятием "популяция".
Популяция - совокупность особей, длительное время обитающих на одной территории. При этом контакты между особями одной популяции происходят чаще, чем между особями разных.
Раз люди одной этнической принадлежности имеют общее происхождение и проживают на одной территории, то следующим логичным предположением становится единая для них культура и язык. При этом, когда мы говорим про культуру, то мы подразумеваем и материальные и духовные её проявления, если не углубляться далеко в древность.
Казалось бы, раз люди одного этноса имеют общее происхождение, совокупляются длительное время преимущественно внутри своей популяции, а соответственно должны иметь и одинаковую культуру и одинаковый язык, то и ДНК у них должна быть условно одинакова и внешний вид схожий.
На это всё можно лишь показать говорящую картинку:
Для начала, в эти стройные логичные измышления вмешивается последний признак, который отделяет один этнос от другого - самосознание.
Самосознание
Самосознание - сознание человеком самого себя в отличие от других людей, сознание принадлежности к какой-либо общности.
Недостаточно иметь одинаковым ранее перечисленные признаки, если совокупность людей разбивается на части, которые осознают себя разными. В принципе, как-то так изначально происходит выделение одного этноса из другого, а потом уже они набирают дополнительные отличия друг от друга. Многие тысячи лет назад, к примеру, постепенно рассыпалась праиндоевропейская общность на разные ветви, разойдясь в итоге на разные языки, разные культуры, и даже кардинально разные внешности - сравните армян и шведов. Это если говорить масштабно.
А если говорить более яркими примерами, то чисто славянский ребёнок, выкраденный цыганами, будет воспитан в их культуре и будет потом осознавать себя цыганом.
Ещё одна говорящая картинка
То есть, если воспитанный в одном этносе человек, будет осознавать себя представителем другого, то ни что ему не может помешать им быть, если остальные члены общности его принимают.
Накидаю немного примеров.
Исидор Ростовский - православный святой русский чудотворец, изначально германский католик западнославянского происхождения.
Владимир Иванович Даль - русский писатель, этнограф, автор знаменитого толкового словаря, сын датчанина и немки.
Список великих русских людей иностранного происхождения можно продолжать достаточно долго. Срача ради можно упомянуть русскую императрицу немецкого происхождения Екатерину II с её "Я немка по рождению и русская душой".
Ференц Лист - композитор австрийского происхождения, родной язык - немецкий. Считается венгерским композитором, идентифицировал себя венгром, на венгерском языке начал говорить ближе к концу жизни.
Бывают и обратные примеры. Поволжские немцы считали себя немцами, и на стыке веков часть из них поехала к своим корням в Германию с надеждой жить "как немцы среди немцев", но в итоге оказались там чужими. Позже некоторые из них осознали: "В России нам доставалось за то, что мы немцы, а тут мы вдруг стали русскими".
Да, здесь возможны "химеры", когда представители негроидной внешности (вспоминаем детей олимпиады) могут считать себя русскими, но "исконно" русские их таковыми считают с натяжкой. Впрочем, у нас достаточно толерантный в этом плане народ.
Ладно, будем пока считать, что самосознание является неким исключительным случаем из признаков этноса. Давайте поговорим об остальных признаках чуть подробней.
Я пока не встречал упоминания этноса, который бы имел более одного "происхождения" - то есть имел бы два или более независимых места-времени зарождения. Пропустим этот признак. Перейдем к территории проживания.
Территория
Определение исходит из того, что для этноса можно очертить некоторую территорию, на котором он проживает. И здесь кроется маленький подвох - эта территория в большинстве случаев не имеет чётких границ. По сути, мы говорим не о строго очерченной местности, а о некоем "облаке", где в условном центре концентрация будет максимальной, а на краях меньше.
Если кто-то считает, что государственную границу можно считать ограничивающим фактором для территории этноса, то пусть поищет бельгийский этнос.
При этом не стоит забывать, что такая популяция обычно не живёт в изоляции. Рядом будет существовать ряд популяций других этносов - и тоже в виде облака. И облака эти будут взаимно перекрываться. Значит, будут существовать территории, общие для более чем одного этноса. А это будет означать, что где-то в местах перекрытия облаков будет происходить активный обмен культурой, языком, генами, т.е. признаки этноса будут размываться.
Ещё в прошлом веке зачастую для всех этносов постулировалась автохтонность. То есть этнос "где родился, там и пригодился". Предполагалось, что территория этноса может меняться только за счет естественного прироста/убыли, а миграции этносов рассматривались как исключительные случаи. Однако более глубокое изучение показало, что миграция этносов не дальние расстояния является вполне обыденным явлением.
Продолжая примеры из нашей праиндоевропейской истории, можно вспомнить, что иранцы оказались на своей текущей территории за счёт нескольких миграций откуда-то из причерноморских степей. Тохары, по предположению Л.С.Клейна, отправились в свою миграцию с северо-запада от тех же степей и пришли на север Афганистана.
Из более близких к нам миграций можно упомянуть массовое переселение волжских калмыков с берегов Волги на территорию Китая в XVIII веке, которое известно как исход в Джунгарию или Торгутский побег. Калмыки выдвинулись в путь в количестве около 150 тысяч человек - со своими семьями и хозяйством, и с боями прошли его по враждебной территории почти за 7 месяцев. (И после этого некоторые будут отрицать возможность монгольского войска дойти до Руси за несколько лет.)
Исход калмыков из России в Джунгарию (Китай) в 1771 году. Гравюра Ш.-М. Жоффруа, 1845
Естественно, что за время миграции, если она не была такая же поспешная, этнос по дороге будет так или иначе взаимодействовать с местным населением, что приведет к смешиванию культуры, языка, генетики. Некоторые этносы при этом переродятся во что-то иное, а некоторые вполне будут считать себя всё теми же "изначальными". Как яркий пример - венгры, которые переселились с Южного Урала в Карпатский бассейн. Они сохранили язык, привнесли на территорию славян свою культуру и существенно потеряли генетику.
В общем, я к тому, что территория не является явным маркером для того, чтобы утверждать что вот на этой земле проживают представители одного этноса, а на той земле представители другого. Это этнос привязан к некоему ареалу в пространстве и времени.
Впрочем, существуют случаи, когда территория популяции ограничена естественными преградами - так получаются народы-изоляты.
Культура
Культура должна бы стать одним из основных признаков различий между этносами. Однако, в силу соседства с другими этносами, да и не просто соседства, а долговременного связанного исторической судьбой, культуры соседних этносов во многом схожи. Так схожи избы южнорусских земель и восточноукраинских, псковских и белорусских.
Попробуйте угадать, какой из орнаментов полотенец какому этносу принадлежит:
(Подсказка - один из них финно-угорский)
Кстати, элементы конкретно этих орнаментов восходят к языческим временам, а ещё точнее к культу Матери-Земли и божественных близнецов. Культ к 19 веку уже остался в поговорках и присказках, а орнамент ещё жив.
В итоге, культура у нас является набором разных элементов, каждый из которых имеет своё собственное размытое распространение по ареалу и собственный центр концентрации. Глядя на любой из элементов культуры, мы не сможем сделать четкий вывод об этносе. Теоретически, имея набор элементов, мы сможем показать территорию, на которых они встречаются все вместе. Однако культура может в себя включать множество таких наборов. Таким образом, говоря об этносе, мы можем выделить наборы элементов культуры, присущие ему.
Ладно. С отсутствием четких границ у территории популяции этноса всё вроде понятно, как и с последующими проблемами в отсутствии границ у культуры. Значит можно опереться на язык!
Язык
В принципе, можно сказать, что раз для меня русский язык является родным, значит я - русский. Хм. А что делать тем, кто с детства учил 2 языка как родные? Ведь это распространенная практика, например, для некоторых народов России. Да и в остальных частях света таких мест вполне предостаточно.
Ладно, будем считать, что люди с более чем одним родным языком могут самоопределиться. А что тогда делать, например, с карелами? К примеру, по переписи 2010 года тверских карел у нас в стране было более 7 тысяч человек. А вот карельским языком из них владели только 25 человек. И то, это определялось на основании опроса.
Хорошо, будем считать, что карелы, да ещё и тверские, это всего лишь какой-то субэтнос.
Давайте взглянем на ирландцев. Согласно переписи, в Ирландии проживает 5 млн. человек. Согласно опросам (я не буду повторно лепить картинку) 1,6 млн. из проживающих там говорит на ирландском языке. Согласно же оценкам лингвистов, лишь 65 тысяч человек использует его в повседневном общении, а всего владеющих они оценивают менее чем в 200 тысяч.
Гэлтахты — районы, где до сих пор сохраняется разговорный ирландский язык в Ирландии.
То есть подавляющее большинство ирландцев знает и использует лишь английский язык. Мы не будем их считать ирландцами?.
В принципе, зная этнос, мы можем сделать предположение о языке его носителя. Обратное верно не всегда.
Но по внешнему виду мы ведь можем отличить испанца от русского!
Внешний вид
Этнографы давно уже собрали данные по внешнему виду - это достаточно простое действие, не требующее сложного оборудования, да и к тому же такие данные собирают уже более сотни лет. Параметров, по которым измеряют внешние отличия, порядка 50. Можно среди них выделить следующие достаточно простые - рост, профильность лица (от монголоидного плоского до высокопрофилированного римского), форма носа и его ширина, расстояние между скулами, цвет глаз и волос, характер папиллярного узора на подушечках пальцев и ладони.
Не буду никого ни в чём уверять, просто предлагаю посмотреть на карте данные по этническим русским.
Средний рост бороды. Темный цвет - максимальный вариант (т.е. третичный волосяной покров доходит почти до глаз)
Длина тела. Темный - выше.
(остальные картинки можно посмотреть в книге "Русский генофонд на Русской равнине", Балановская Е. В., Балановский О. П.)
Если составить обобщенную антропологическую карту для русских, то мы и там увидим ареалы отклонений.
С другой стороны, если взять за популяцию уже не русский этнос, а более высокий уровень - восточноевропейский, то на его фоне отклонения будут уже минимальны.
Обобщённая карта антропологического состава Восточной Европы.
Получается, что по внешнему виду нельзя сделать однозначный вывод об этническом происхождении. Впрочем, по принадлежности к этносу можно сделать предположения о внешнем виде. Усредненном. Очень усредненном внешнем виде.
Раз поговорили про внешний вид, то можно перейти и к генетике.
Генетика
Каждый из нас является продуктом своих родителей. Мы содержим гены как отца, так и матери, которые передаются случайным смешением. Но есть и небольшие исключения. Y-хромосома передаётся исключительно от отца к сыну. Есть еще гены, которые у человека передаются от матери к детям - это внеядерная мтДНК. Из-за этого мужчины получают "преимущество" - у них есть отличительная часть как от отца, так и от матери. Разные мутации этих ДНК разделили на гаплогруппы и проследили их изменения (мутации), выстроив деревья предковых вариантов.
Имеются данные, что всё же возможна передача "отцовской" мтДНК в очень исключительных случаях.
Ещё каких-то десять лет назад было популярно изучать именно эти две части генетики, так как они давали картину по линии отцовства и по линии материнства. Ведь эти гены могут давать представление о предках.
Однако тут кроется и "небольшая" проблема - ни одна из этих ДНК не даёт полную историческую картину по предкам. Например, мы не увидим тут дедушку по материнской линии или бабушку по отцовской. А на уровне прапра-, к примеру, мы вообще увидим только 2х человек из 16.
Примерно 10 лет назад начали использовать полногеномный анализ, который позволяет составить полную картину по генам популяций.
Полногеномный анализ дал новый толчок исследованиям настоящего и прошлого нашего человечества. Пример такого исследования по родственным бракам можно обнаружить в статье Инцест. Историческая действительность
В результате выяснилось... В общем, выяснилось примерно то же, что и по внешности - полный разброд и шатание.
Для человечества примерно 15 % генетической изменчивости соответствует различиям между всеми популяциями мира, а примерно 85 % — приходится на различия между организмами.
При этом ещё стоит добавить, что гены подвержены естественному отбору - люди с подходящими генами для выживания на конкретной территории будут передавать свои гены следующему поколению чаще. А значит, что срезы генофонда этноса в разные периоды времени будут отличаться друг от друга даже в условиях полной изоляции.
Привязать какие-либо гены к народу - невозможно. Ген (аллели гена) может "возникнуть" в одном месте, а "размножиться" совершенно в другом, далёком от изначального, за счёт миграций. Люди, как носители конкретных генов, вольны в выборе "родного" этноса. Популяции, как общности людей, существуют в условиях взаимопроникновения с другими популяциями и обмениваются с ними генами.
В итоге, по генетике конкретного человека мы не можем определить его этнос. Говоря об этносе, мы можем дать расклад по генофонду, то есть встречаемости различных аллелей генов в его популяции в некоторый момент времени.
Используя единую базу генетических и лингвистических данных для 397 популяций, говорящих на 295 языках, специалисты провели глобальный анализ генетически-языковых соответствий и несоответствий в населении мира. Примерно в 18% случаев обнаружились несоответствия. Подробнее здесь.
Этнические тесты ДНК
А как же этнические тесты ДНК, которые рекламируют лаборатории?
А никак. Это чистое шарлатанство с использованием слова "этнос".
На самом деле, по итогам такого теста при правильно проведенном анализе можно говорить о близости к различным популяциям людей, но не об этничности.
Итак, а что же такое этнос и как же его определить? Накидывайте варианты в комментариях :)
О, остался же нерассмотренным вопрос про идентичность ДНК сиблингов.
Если говорить именно про тест на "этническую принадлежность", то результаты у них будут одинаковыми в случае однояйцевых близнецов. В общем же случае результат будет неидентичен, но скорее всего достаточно схож.
Кстати, а вы знали, что однояйцевые близнецы в большинстве случаев отличаются по ДНК?